
前頭側頭葉変性症原因遺伝子VCPはポリグルタミン病の共通病態を制御する (2013)
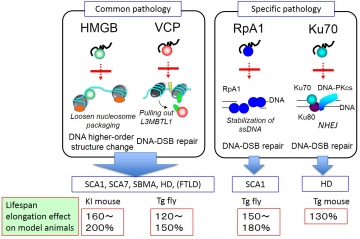
私たちの研究室は、アルツハイマー病、パーキンソン病に次いで頻度の高い神経変性疾患であるポリグルタミン病の病態解明に取り組んでいるが、これらの変性疾患の原因タンパク質は正常タンパク質と結合して機能阻害を起こすと考えられている。私たちは10数年前に、ポリグルタミン配列に結合する新規タンパク質をyeast two-hybrid法により検索し、PQBP1と共に発見した分子がTERA/VCPである (Imafuku et al., BBRC 1998)。その後、他のグループによりVCPのポリグルタミン病タンパク質の一種(Ataxin 3)への結合と病態への関与が再検証されていた(Hirabayashi et al., Cell Death & Differ. 2001)。しかしながら、VCPがどのような分子機構でポリグルタミン病態を制御するのか、他のポリグルタミン病タンパク質にも影響を与えるのかなど解明しなければならない点が数多く残されていた。VCPはAAA ATPaseファミリーに属するタンパクであるが、「膜輸送」「小胞体関連タンパク質分解」「DNA損傷修復」などの様々な細胞機能においてその活性が必要である。また、VCP遺伝子変異自体も前頭側頭葉変性症の原因となることが近年明らかとなっている。
これらの現状を踏まえると、ポリグルタミン病においてはVCPを介した共通病態が存在するという仮説が可能になる。また、変異型ポリグルタミン病タンパク質がVCPをトラップすることで神経変性に至るとすれば、多岐にわたるVCP機能のどれが阻害されることが重要であるろうか?本年度の研究において、まずはVCPが実際に様々なポリグルタミン病タンパク質と結合するかを検討した。Ataxin-1(脊髄小脳失調症1型(SCA1)の原因遺伝子)、Ataxin-7(脊髄小脳失調症7型の原因遺伝子)、アンドロジェン受容体(球脊髄性筋萎縮症の原因遺伝子)、ハンチンチン(ハンチントン病(HD)の原因遺伝子)という、4種類のポリグルタミン病の疾患遺伝子を用いて検討したところ、正常型、変異型ともにポリグルタミン病タンパク質はVCPに結合し、ポリグルタミン病タンパク質からポリグルタミン配列だけを除いた変異体タンパク質とは結合しなかった。これにより、VCPとポリグルタミン病タンパク質の関係が一般化された。そこで、VCP神経細胞内のどこで働くか、正常マウスの細胞内局在を検討した。VCPはプルキンエ細胞及び脊髄運動ニューロンでは核に強く局在し、神経細胞においてVCPが核での機能を発揮していることが示唆された。私たちはすでに、ポリグルタミン病における核DNAの二重鎖切断亢進という病態を見出しており(Enokido et al., JCB 2010)、VCPの阻害がDNAダメージの修復不全を介して病態に関与するのではないかと考え更なる実験を行った。
HDおよびSCA1モデルマウスとモデルショウジョウバエにおいて、DNA二重鎖切断のマーカーであるリン酸化ヒストン(gammaH2AX及びgammaH2Av)の免疫染色を行ったところ、これらのモデルではいずれもDNA二重鎖切断の亢進が確認された。さらに、モデルショウジョウバエにおいてVCPを過剰発現させたところ、寿命の短縮といった症状のみならず、gammaH2Avシグナルの低下も顕著であった。VCPはDNA損傷部位に集積することでDNA二重鎖切断を促進する機能を持つ。そこで、マイクロレーザー照射法を用いて解析したところ、変異型ハンチンチン及びAtaxin-1を発現する細胞ではVCPのダメージ部位への集積が阻害されていた。
これらの結果は、1)ポリグルタミン病タンパク質はその種類によらず、TERA/VCP/p97と結合すること、2)変異ポリグルタミン病タンパク質がVCPと結合した際にVCPの細胞内動態が変化するために、DNA損傷部位への移動を妨げてDNA修復能が低下し、最終的にDNA二重鎖切断が増加すること、を示している。近年、DNA二重鎖切断は正常な神経活動によっても誘導されることが明らかになった(Suberbielle et al., Nat Neurosci 2013)が、このように非病態下で通常に生じるDNA二重鎖切断に対してVCPは修復を行っており、変異ポリグルタミン病タンパク質がこの機能を阻害すると考えられる。現在、ウィルスベクターによるモデルマウスの治療を目指しており、今後の治療応用が期待される。
これらの結果は、1)ポリグルタミン病タンパク質はその種類によらず、TERA/VCP/p97と結合すること、2)変異ポリグルタミン病タンパク質がVCPと結合した際にVCPの細胞内動態が変化するために、DNA損傷部位への移動を妨げてDNA修復能が低下し、最終的にDNA二重鎖切断が増加すること、を示している。近年、DNA二重鎖切断は正常な神経活動によっても誘導されることが明らかになった(Suberbielle et al., Nat Neurosci 2013)が、このように非病態下で通常に生じるDNA二重鎖切断に対してVCPは修復を行っており、変異ポリグルタミン病タンパク質がこの機能を阻害すると考えられる。現在、ウィルスベクターによるモデルマウスの治療を目指しており、今後の治療応用が期待される。
発表論文
Fujita, K., Nakamura, Y., Oka, T., Ito, H., Tamura, T., Tagawa, K., Sasabe, T., Katsuta, A., Motoki, K., Shiwaku, H., Sone, M., Yoshida, C., Katsuno, M., Eishi, Y., Murata, M., Taylor, JP., Wanker, EE., Kono, K., Tashiro, S., Sobue, G., La, Spada, AR., and Okazawa, H.
A functional deficiency of TERA/VCP/p97 contributes to impaired DNA damage repair in multiple polyglutamine diseases. Nature Commun. 4:1816. doi: 10.1038/ncomms2828
A functional deficiency of TERA/VCP/p97 contributes to impaired DNA damage repair in multiple polyglutamine diseases. Nature Commun. 4:1816. doi: 10.1038/ncomms2828