
「サル免疫不全ウイルスのタンパク質がヒトの抗ウイルスタンパク質BST-2 の機能を妨害するメカニズムの解明」【山岡昇司 教授、芳田剛 助教】
公開日:2022.2.24
「 サル免疫不全ウイルスのタンパク質がヒトの抗ウイルスタンパク質BST-2
の機能を妨害するメカニズムの解明 」
― 動物に感染するウイルスが宿主域を拡大し、ヒトへ伝染するメカニズムの解明へ ―
の機能を妨害するメカニズムの解明 」
― 動物に感染するウイルスが宿主域を拡大し、ヒトへ伝染するメカニズムの解明へ ―
ポイント
- アフリカに生息するサルから単離されたサル免疫不全ウイルスのタンパク質のアミノ酸で、ヒトの防御タンパク質BST-2を妨害する上で重要なものを同定しました。
- それら同定されたアミノ酸が、自然宿主サルのBST-2を妨害する上で重要なアミノ酸とは異なることがわかり、サルに加えヒトに感染するための機能をウイルスが獲得していた可能性が示されました。
- 本研究は、現在動物に感染しているウイルスの「ヒトへの伝播リスクの評価」に寄与し、今後起こり 得る新規感染症発生の未然予測へ貢献するものと考えられます。
- ヒトとサルのBST-2は妨害されるメカニズムがそれぞれ異なることから、動物も「防御に関わる遺伝子」を進化させている可能性が示されました。
東京医科歯科大学大学院医歯学総合研究科ウイルス制御学分野の芳田剛助教と山岡昇司教授の研究グループは、アメリカ国立衛生研究所(NIH)との共同研究で、野生動物から分離されたウイルスのタンパク質がヒトの防御機構を妨害する分子メカニズムを明らかにしました。この研究は文部科学省科学研究費補助金ならびに一般財団法人中辻創智社等の支援のもとでおこなわれたもので、その研究成果は、国際科学誌Journal of Virologyに、2022年2月23日にオンライン版で発表されました。
研究の背景
新型コロナウイルスをはじめ、ヒトに感染症を起こすウイルスの多くは、他の動物に感染しているウイルスがヒトへ伝染してきたものであると考えられています。実際、エイズの原因ウイルスであるヒト免疫不全ウイルス1型(HIV-1、※①)も、アフリカに生息するサルに感染する免疫不全ウイルス(SIV、※②)がチンパンジーに伝染した後、さらにヒトへ伝染したものであると考えられています。ウイルスが新たな動物種に伝染するには、当然のことながら、伝播先の動物の防御機構を超えなければなりません。
動物のタンパク質BST-2(別名Tetherin、※③) は、ウイルス感染に対して防御的に働くタンパク質の1つで、ウイルスの放出を抑制する機能を持っています。しかし、HIV-1は ウイルスタンパク質Vpu(※④)を用いてヒトのBST-2機能を妨害し、BST-2に打ち勝つことで増殖することが知られています。しかし、これまでに <ヒト 以外の動物に感染するウイルス> がヒトBST-2を妨害する例は報告されていませんでした。
動物のタンパク質BST-2(別名Tetherin、※③) は、ウイルス感染に対して防御的に働くタンパク質の1つで、ウイルスの放出を抑制する機能を持っています。しかし、HIV-1は ウイルスタンパク質Vpu(※④)を用いてヒトのBST-2機能を妨害し、BST-2に打ち勝つことで増殖することが知られています。しかし、これまでに <ヒト 以外の動物に感染するウイルス> がヒトBST-2を妨害する例は報告されていませんでした。
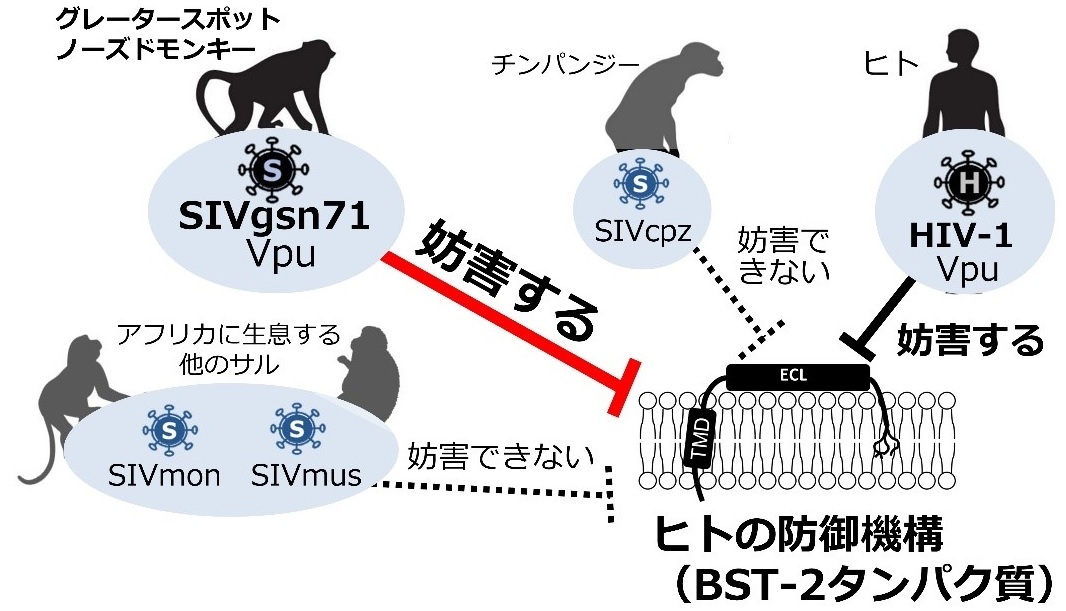
図-1 「動物に感染するウイルス」によるヒト防御機構の妨害
研究成果の概要
2020年に研究グループは、アフリカに生息するグレータースポットノーズドモンキー(greater spot-nosed monkey:以降サルと呼称)から単離されたウイルスSIVgsnの99CM71株のVpuタンパク質がヒトのBST-2の機能を妨害することを報告し(Yao and Yoshida et al., Journal of Virology、2020)、<野生動物に感染するウイルス>がヒトの防御機構を克服できる術をすでに獲得している危険性について、警鐘を鳴らしてきました(図-1)。
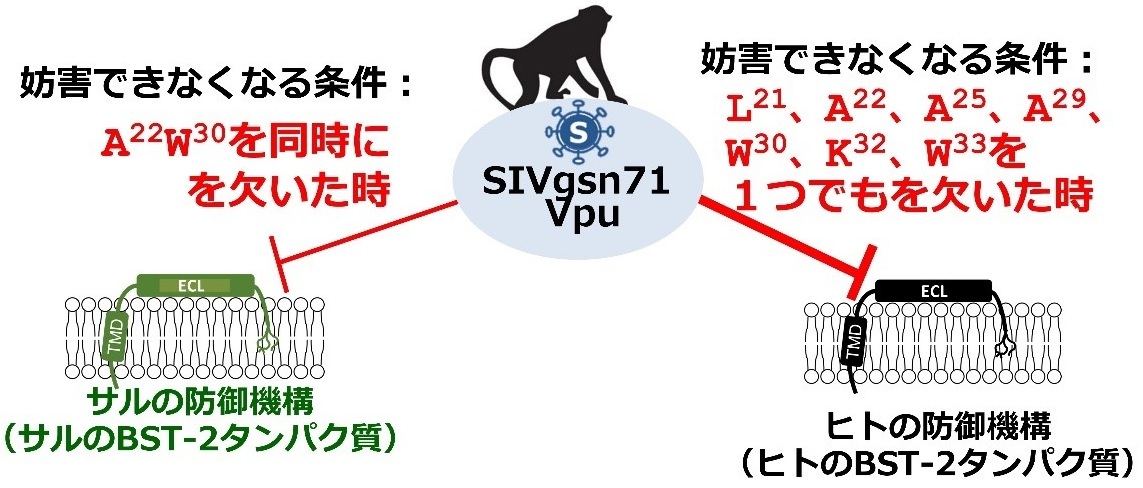
今回、研究グループは、SIVgsnのVpuタンパク質を構成するアミノ酸のうち、ヒトのBST-2機能を妨害する上で重要な役割を担う7個のアミノ酸を同定しました。この7アミノ酸のうちの1つでも欠けてしまうと(1つに置換変異を加えるだけで)、ヒトBST-2妨害機能を失うことがわかりました。一方で、このVpuタンパク質の サルBST-2を妨害する機能を失わせるには複数のアミノ酸を同時に置換する必要があり、1つのアミノ酸の置換変異によりサルBST-2妨害機能を失わせることはできませんでした(図-2)。これらの結果から、SIVgsnの99CM71株のVpuタンパク質は、それぞれサルとヒトのBST-2を異なるメカニズムにより妨害していることがわかりました。
今回、研究グループは、SIVgsnのVpuタンパク質を構成するアミノ酸のうち、ヒトのBST-2機能を妨害する上で重要な役割を担う7個のアミノ酸を同定しました。この7アミノ酸のうちの1つでも欠けてしまうと(1つに置換変異を加えるだけで)、ヒトBST-2妨害機能を失うことがわかりました。一方で、このVpuタンパク質の サルBST-2を妨害する機能を失わせるには複数のアミノ酸を同時に置換する必要があり、1つのアミノ酸の置換変異によりサルBST-2妨害機能を失わせることはできませんでした(図-2)。これらの結果から、SIVgsnの99CM71株のVpuタンパク質は、それぞれサルとヒトのBST-2を異なるメカニズムにより妨害していることがわかりました。
図-2:このウイルスタンパク質はヒトとサルの防御機構を異なるメカニズムで妨害していた
研究成果の意義
野生動物に感染しているウイルスが、現在感染している宿主動物の防御機構を妨害する機能とは異なる、他の動物(ヒト)の防御機構を妨害する機能を持っていることが判明し、ウイルスが潜在的に他の動物へ伝染するための準備をしている可能性が示唆されました。ヒトのBST-2を妨害する機能を獲得することが、<動物に感染するウイルス> がヒトへ伝染するための1つの必要条件である可能性があるため、本研究が「動物に感染しているウイルスのヒトへの伝播リスク評価」に寄与し、今後起こり得る新規感染症発生の未然予測へ貢献するものと考えられます。
一方で、ウイルスに対する動物の防御タンパク質に着目しながらこの結果を見ると、ヒトとサルのBST-2は妨害される分子機序が異なることが判明したことになります。この事実は、動物も進化の中で「防御に関わる遺伝子」を変化させている可能性を示唆しています。ウイルスに対する防御は動物の生存と密接に関連しているため、BST-2の機能、そしてそれを規定する遺伝子の配列が、進化の歴史の中で淘汰(絶滅)と自然選択(生存)の分水嶺となってきた可能性が示されました。
一方で、ウイルスに対する動物の防御タンパク質に着目しながらこの結果を見ると、ヒトとサルのBST-2は妨害される分子機序が異なることが判明したことになります。この事実は、動物も進化の中で「防御に関わる遺伝子」を変化させている可能性を示唆しています。ウイルスに対する防御は動物の生存と密接に関連しているため、BST-2の機能、そしてそれを規定する遺伝子の配列が、進化の歴史の中で淘汰(絶滅)と自然選択(生存)の分水嶺となってきた可能性が示されました。
用語解説
※①ヒト免疫不全ウイルス1型(HIV-1)・・・・・・・・後天性免疫不全症候群(エイズ)の原因ウイルスです。チンパンジーに感染するウイルスがヒトに伝染したことを機に、150年前頃に発生したと考えられています。
※②サル免疫不全ウイルス(SIV)・・・・・・・・SIVの後の小文字3文字は、ウイルスが単離された霊長類動物の名前の略称をあらわす決まりがあります(例えば、チンパンジーから単離されたウイルスはSIVcpzと命名され、グレータースポットノーズドモンキーから単離されたウイルスはSIVgsnと名付けられています)。
※③BST-2・・・・・・・・bone marrow stromal antigen 2。別名Tetherin、CD317。HIV-1以外の種々のウイルス(エンベロープタンパク質を持つウイルス群)に対して、ウイルス放出抑制効果を発揮することが知られています。ウイルス感染により誘導されるタンパク質インターフェロンに依存して発現することも知られています。
※④Vpuタンパク質・・・・・・・・ウイルスが発現する膜タンパク質です。ウイルスによりタンパク質の長さが異なりますが、77から86アミノ酸の長さであり、小さなタンパク質であることが知られています。HIV-1はこのVpuタンパク質を発現しますが、サル免疫不全ウイルス(SIV)には発現するウイルス株(SIVgsn, SIVmon, SIVmus等)と発現しない株(SIVrcm等)の両方が存在します。
※②サル免疫不全ウイルス(SIV)・・・・・・・・SIVの後の小文字3文字は、ウイルスが単離された霊長類動物の名前の略称をあらわす決まりがあります(例えば、チンパンジーから単離されたウイルスはSIVcpzと命名され、グレータースポットノーズドモンキーから単離されたウイルスはSIVgsnと名付けられています)。
※③BST-2・・・・・・・・bone marrow stromal antigen 2。別名Tetherin、CD317。HIV-1以外の種々のウイルス(エンベロープタンパク質を持つウイルス群)に対して、ウイルス放出抑制効果を発揮することが知られています。ウイルス感染により誘導されるタンパク質インターフェロンに依存して発現することも知られています。
※④Vpuタンパク質・・・・・・・・ウイルスが発現する膜タンパク質です。ウイルスによりタンパク質の長さが異なりますが、77から86アミノ酸の長さであり、小さなタンパク質であることが知られています。HIV-1はこのVpuタンパク質を発現しますが、サル免疫不全ウイルス(SIV)には発現するウイルス株(SIVgsn, SIVmon, SIVmus等)と発現しない株(SIVrcm等)の両方が存在します。
論文情報
掲載誌:Journal of Virology
論文タイトル: Simian Immunodeficiency Virus SIVgsn-99CM71 Vpu Employs Different Amino Acids To Antagonize Human and Greater Spot-Nosed Monkey BST-2
DOI:https://doi.org/10.1128/JVI.01527-21
研究者プロフィール

芳田 剛 (ヨシダ タケシ) Takeshi Yoshida
東京医科歯科大学 大学院医歯学総合研究科
ウイルス制御学分野 助教
・研究領域 ウイルス学
進化生物学
霊長類学
東京医科歯科大学 大学院医歯学総合研究科
ウイルス制御学分野 助教
・研究領域 ウイルス学
進化生物学
霊長類学

山岡 昇司(ヤマオカ ショウジ)Shoji Yamaoka
東京医科歯科大学 大学院医歯学総合研究科
教授
・研究領域 ウイルス学
東京医科歯科大学 大学院医歯学総合研究科
教授
・研究領域 ウイルス学
問い合わせ先
<研究に関すること>
東京医科歯科大学大学院医歯学総合研究科
ウイルス制御学分野 芳田 剛(ヨシダ タケシ)
E-mail: takeshi-yoshida[@]umin.ac.jp (芳田)
<報道に関すること>
東京医科歯科大学 総務部総務秘書課広報係
〒113-8510 東京都文京区湯島1-5-45
E-mail:kouhou.adm[@]tmd.ac.jp