
核小体の構造形成の仕組みを解明 (2023)
核小体(「仁」とも呼ぶ)は、細胞核の中でもっとも大きい構造物である。核小体の存在は、光学顕微鏡が発明されていた1830年代には、少なくとも複数の科学者によって記載されており、一説によれば、1770年代に発見されたとも言われている。現在では、さまざまな新しい顕微鏡が開発され、その構造を見ることができるようになってきた。核は膜を持っていて、それ以外の細胞部分と区切られているが、核小体には膜がない。したがって、どのようなメカニズムが核小体の構造を形成・維持するかは長いあいだナゾであった。
当研究グループは、25年ほど前(1998年)に、複数のポリグルタミン病原因タンパク質に共通して含有されるポリグルタミン配列への結合蛋白質をyeast two hybrid法を用いて探索し、PQBP1, 2, 3, 4, 5という5つの新規分子を発見した(Imafuku et al, BBRC 1998; Waragai et al, Hum Mol Genet 1999; Okazawa et al, Neuron 2002)。このうち、PQBP1については、当研究グループに加えて多数の研究者が参入して、基本的な分子機能と、神経変性疾患、小頭症、発達障害、ウイルス感染症、免疫疾患などにおける病態機能が多数報告されている。また、PQBP1はヒトの知的障害の原因遺伝子であることから(Kalscheuer et al, Nature Genet 2003)、ヒト知能に関わる分子であることも確実である。一方、PQBP5を含む、PQBP1以外のPQBPsについては、この25年間にPQBP5などが核小体タンパク質であることが分かったものの(そのためPQBP5にNOL10という新しい名称がつけられた)、十分な知識は得られていなかった。今回はPQBP5に焦点を当てた20年近い研究の末に、当初予測していなかった結論に至った。
今回の研究では、PQBP5がPQBP1と同様に天然変性タンパク質であることが研究進展の契機となった。タンパク質は決まった形の3次元構造(高次構造)を保っているというのが古典的な考え方であった。しかしながら、かなりの割合のタンパク質が、その一部あるいは全体において、安定した高次構造を持たない状態(天然変性状態)にあることが明らかになってきた。しかも、このような天然変性タンパク質の多くが、神経変性疾患の原因タンパク質そのものであったり、原因タンパク質と結合関係にあるタンパク質であったりすることが分かってきた。今回の研究では、予測コンピュータプログラムからPQBP5が天然変性タンパク質である可能性が高まったため、さらに金沢大学の安藤教授らと共同研究を行い、高速原子間力顕微鏡を用いて1分子の挙動を観察した結果、PQBP5が天然変性タンパク質であることを確認した。
天然変性タンパク質には、「液液相分離(LLPS)」という、水と油のように互いの液相と液相が分離する性質があるために、それ以外のタンパク質から分離して液滴(液体状態での集合体)を形成する。核小体タンパク質に天然変性タンパク質が多いことが近年になって解明されたことから、膜を持たない核小体の形成は、このような液滴であり、LLPSの物理的エネルギー安定性が基盤にあるものと考えられるようになった。さらに、他の核小体タンパク質であるfibrillarin, nucleolinと生物物理学的性質を比較することで、PQBP5が他の核小体タンパク質よりも安定した分子であり、PQBP5によってLLPSに基づく集合体(液滴)である核小体が浸透圧や熱などのストレスに強くなり、構造が安定化することを明らかにした。
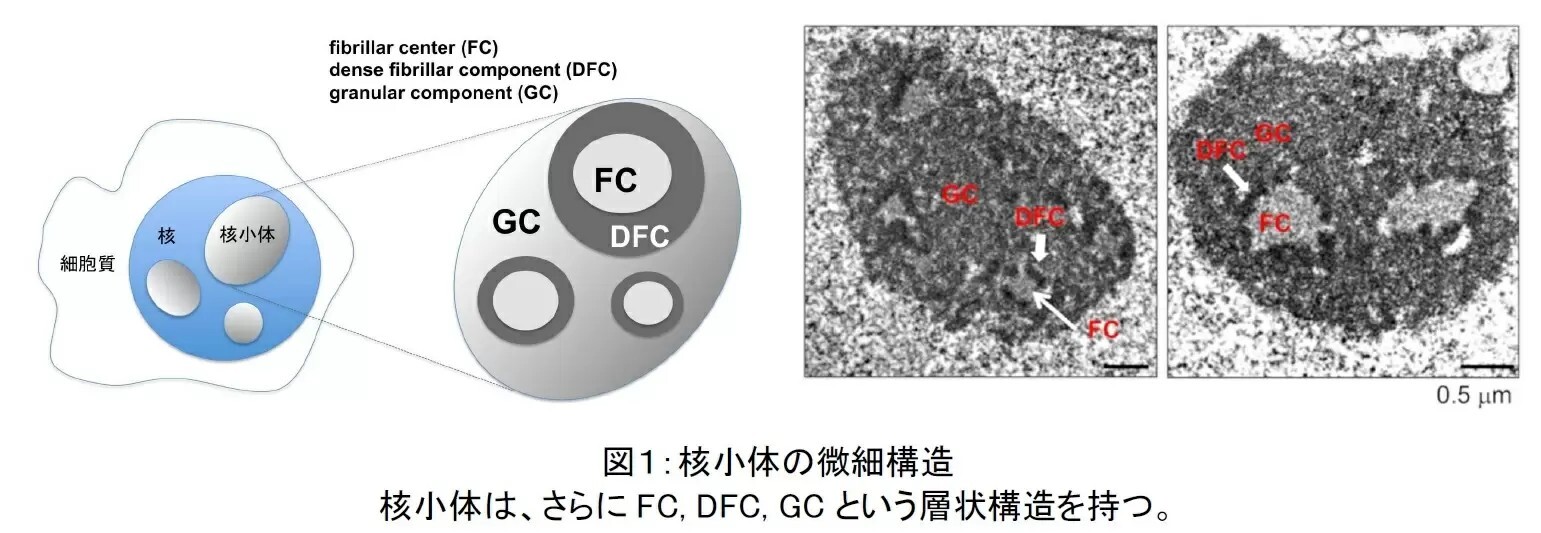
電子顕微鏡で核小体の細部構造を観察すると、核小体にはさらにFC, DFC, GCという細かな構造があることがこれまでに知られている(図1)。
当研究グループは、25年ほど前(1998年)に、複数のポリグルタミン病原因タンパク質に共通して含有されるポリグルタミン配列への結合蛋白質をyeast two hybrid法を用いて探索し、PQBP1, 2, 3, 4, 5という5つの新規分子を発見した(Imafuku et al, BBRC 1998; Waragai et al, Hum Mol Genet 1999; Okazawa et al, Neuron 2002)。このうち、PQBP1については、当研究グループに加えて多数の研究者が参入して、基本的な分子機能と、神経変性疾患、小頭症、発達障害、ウイルス感染症、免疫疾患などにおける病態機能が多数報告されている。また、PQBP1はヒトの知的障害の原因遺伝子であることから(Kalscheuer et al, Nature Genet 2003)、ヒト知能に関わる分子であることも確実である。一方、PQBP5を含む、PQBP1以外のPQBPsについては、この25年間にPQBP5などが核小体タンパク質であることが分かったものの(そのためPQBP5にNOL10という新しい名称がつけられた)、十分な知識は得られていなかった。今回はPQBP5に焦点を当てた20年近い研究の末に、当初予測していなかった結論に至った。
今回の研究では、PQBP5がPQBP1と同様に天然変性タンパク質であることが研究進展の契機となった。タンパク質は決まった形の3次元構造(高次構造)を保っているというのが古典的な考え方であった。しかしながら、かなりの割合のタンパク質が、その一部あるいは全体において、安定した高次構造を持たない状態(天然変性状態)にあることが明らかになってきた。しかも、このような天然変性タンパク質の多くが、神経変性疾患の原因タンパク質そのものであったり、原因タンパク質と結合関係にあるタンパク質であったりすることが分かってきた。今回の研究では、予測コンピュータプログラムからPQBP5が天然変性タンパク質である可能性が高まったため、さらに金沢大学の安藤教授らと共同研究を行い、高速原子間力顕微鏡を用いて1分子の挙動を観察した結果、PQBP5が天然変性タンパク質であることを確認した。
天然変性タンパク質には、「液液相分離(LLPS)」という、水と油のように互いの液相と液相が分離する性質があるために、それ以外のタンパク質から分離して液滴(液体状態での集合体)を形成する。核小体タンパク質に天然変性タンパク質が多いことが近年になって解明されたことから、膜を持たない核小体の形成は、このような液滴であり、LLPSの物理的エネルギー安定性が基盤にあるものと考えられるようになった。さらに、他の核小体タンパク質であるfibrillarin, nucleolinと生物物理学的性質を比較することで、PQBP5が他の核小体タンパク質よりも安定した分子であり、PQBP5によってLLPSに基づく集合体(液滴)である核小体が浸透圧や熱などのストレスに強くなり、構造が安定化することを明らかにした。
電子顕微鏡で核小体の細部構造を観察すると、核小体にはさらにFC, DFC, GCという細かな構造があることがこれまでに知られている(図1)。
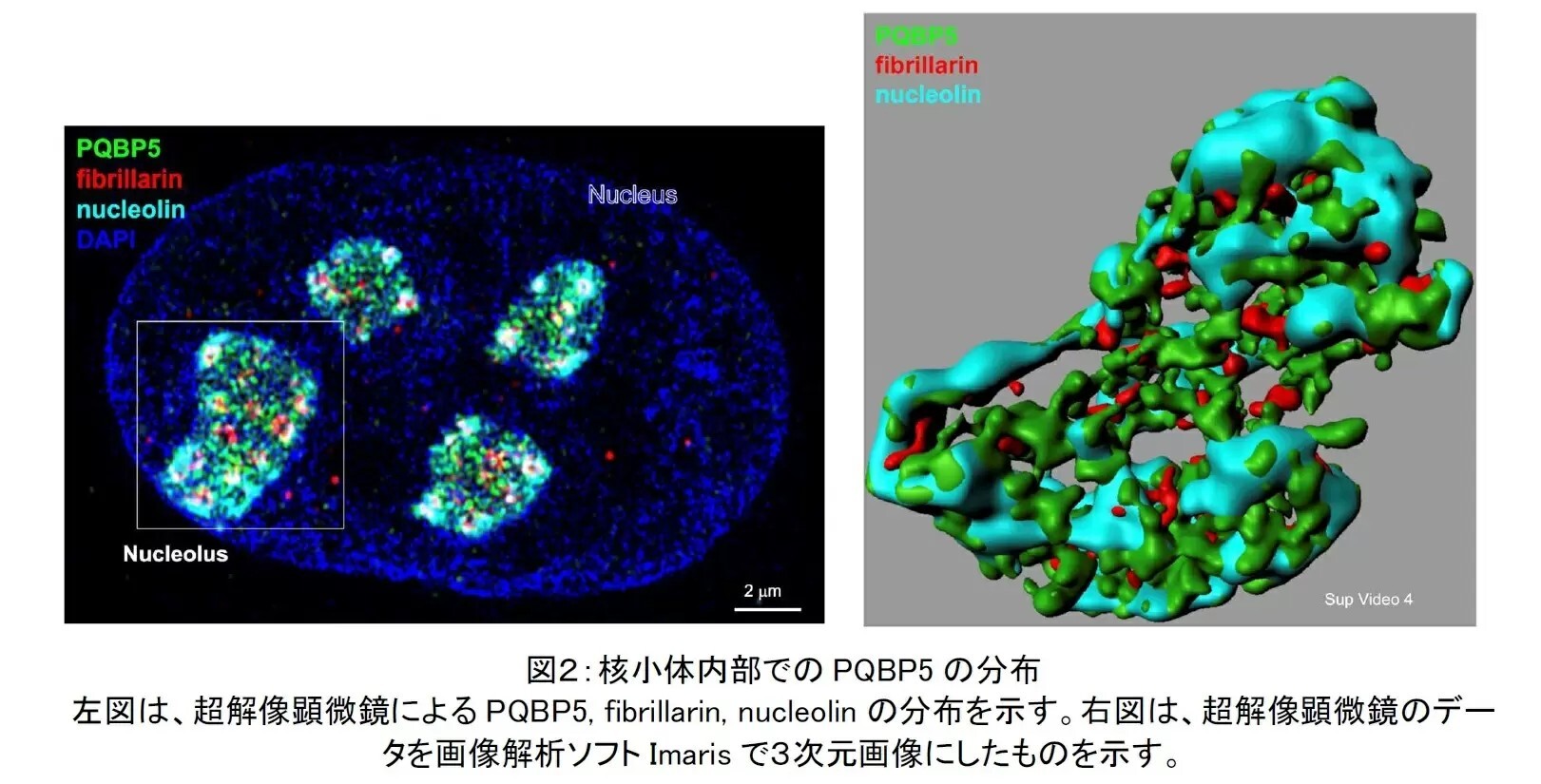
今回の研究では、超解像顕微鏡と CLEM (Correlative light and electron microscopy)という最新技術を用いて、PQBP5の核小体内部における分布を調べた結果、PQBP5は主にGCに存在し、竹で編んだ籠のような構造(籠状構造)を構築して、DFCに分布するfibrillarinや核小体の外皮に存在するnucleolinなどが集合するための骨組みとなっていることを解明した(図2)。 実際に、PQBP5をノックダウンすると、核小体が変形したり消失したりすることも確認した。
さらに、25年前の疑問に戻って、PQBP5と神経変性疾患との関連についても検討した。ハンチントン病の原因タンパク質であるハンチンチンと脊髄小脳失調症1型の原因タンパク質であるアタキシン1は、正常型と異常型は共にPQBP5と結合するものの、PQBP5は異常型の封入体に取り込まれて、核小体から不足することが示された。これは、前述のPQBP5ノックダウンと同様な病態であり、実際、2つの疾患のモデルマウスにおいて、細胞のPQBP5ノックダウンと同様な核小体異常が検出された。
さらに、25年前の疑問に戻って、PQBP5と神経変性疾患との関連についても検討した。ハンチントン病の原因タンパク質であるハンチンチンと脊髄小脳失調症1型の原因タンパク質であるアタキシン1は、正常型と異常型は共にPQBP5と結合するものの、PQBP5は異常型の封入体に取り込まれて、核小体から不足することが示された。これは、前述のPQBP5ノックダウンと同様な病態であり、実際、2つの疾患のモデルマウスにおいて、細胞のPQBP5ノックダウンと同様な核小体異常が検出された。
今回の研究では、1) 核小体が天然変性タンパク質の生物物理的性質を基盤とした複数の構成分子の集合体であることを解明した。 加えて、2) 構成分子の生物物理的性質には(当然ながら)差があり、もっとも安定なPQBP5を基軸に核小体が形成されていること、 3) 浸透圧変化などの細胞ストレス化では核小体構造が乱れるが、PQBP5は集合体として定位置に残り、細胞ストレスからの回復過程において、アンカーとして他の核小体分子の再集合に寄与すること、を示した。
核小体はribosomal DNAからribosomal RNAが産生される場所で形成されると考えられる。PQBP5のLLPS集合体形成もribosomal RNAによって促進することを今回確認しており、この考え方と合致した結果であった。一方、PQBP5ノックダウンはribosomal RNAを最終的に減少させるが、PQBP5タンパク質が減少し、ribosomal RNAが未だ減少していない時点で、すでに核小体構造が崩れていることから、ribosomal RNAが産生されても、PQBP5タンパク質がなければ、核小体は形成できない、と考えられる。
また、今回の研究では、4) 神経変性疾患の原因タンパク質が細胞内凝集体(封入体)を作る過程でPQBP5を巻き込んで(sequestration仮説)、核小体におけるPQBP5を減少させるために、核小体そのものの構造異常を起こすことを示した。今回の研究では、脊髄小脳失調症、ハンチントン病や球脊髄性筋萎縮症(Kennedy病)を含むポリグルタミン病と呼ばれる神経変性疾患グループにおいて、PQBP5の巻き込みによる核小体病態が実際に生じていることを示した。
ポリグルタミン病以外に、どのような神経変性疾患において、同様にPQBP5を介した核小体病態が起きているのかは(例えば、アルツハイマー病やパーキンソン病でも同一病態が起きているかは)、PQBP5に結合しうる変性疾患原因タンパク質の種類が網羅的に確認されていないために、現時点では不明である。しかし、今後さらに研究を続けることができれば、種々の神経疾患における核小体病態の広がりを包括的に捉えることが可能となるであろう。
さらに、PQBP5遺伝子は、がん、老化などにも影響を与えていることが予想される。核小体の大きさ・数が、がん細胞の病理学的診断の根拠に使われているように、核小体は細胞増殖活性と関連している。脳以外の全身臓器では細胞増殖が低下することをセネセンスと呼び、老化の現象の一つと考えられている。これまでに、がんとPQBP5の遺伝子変異や発現量が関係するという論文が複数発表されているが、PQBP5が核小体を守る機能があること、すなわち細胞増殖能力や抗がん剤耐性へ影響しうることと関係があるのではとも予想される。今後、PQBP5の研究が進むことで、がんや老化の制御につながる可能性もある。
核小体はribosomal DNAからribosomal RNAが産生される場所で形成されると考えられる。PQBP5のLLPS集合体形成もribosomal RNAによって促進することを今回確認しており、この考え方と合致した結果であった。一方、PQBP5ノックダウンはribosomal RNAを最終的に減少させるが、PQBP5タンパク質が減少し、ribosomal RNAが未だ減少していない時点で、すでに核小体構造が崩れていることから、ribosomal RNAが産生されても、PQBP5タンパク質がなければ、核小体は形成できない、と考えられる。
また、今回の研究では、4) 神経変性疾患の原因タンパク質が細胞内凝集体(封入体)を作る過程でPQBP5を巻き込んで(sequestration仮説)、核小体におけるPQBP5を減少させるために、核小体そのものの構造異常を起こすことを示した。今回の研究では、脊髄小脳失調症、ハンチントン病や球脊髄性筋萎縮症(Kennedy病)を含むポリグルタミン病と呼ばれる神経変性疾患グループにおいて、PQBP5の巻き込みによる核小体病態が実際に生じていることを示した。
ポリグルタミン病以外に、どのような神経変性疾患において、同様にPQBP5を介した核小体病態が起きているのかは(例えば、アルツハイマー病やパーキンソン病でも同一病態が起きているかは)、PQBP5に結合しうる変性疾患原因タンパク質の種類が網羅的に確認されていないために、現時点では不明である。しかし、今後さらに研究を続けることができれば、種々の神経疾患における核小体病態の広がりを包括的に捉えることが可能となるであろう。
さらに、PQBP5遺伝子は、がん、老化などにも影響を与えていることが予想される。核小体の大きさ・数が、がん細胞の病理学的診断の根拠に使われているように、核小体は細胞増殖活性と関連している。脳以外の全身臓器では細胞増殖が低下することをセネセンスと呼び、老化の現象の一つと考えられている。これまでに、がんとPQBP5の遺伝子変異や発現量が関係するという論文が複数発表されているが、PQBP5が核小体を守る機能があること、すなわち細胞増殖能力や抗がん剤耐性へ影響しうることと関係があるのではとも予想される。今後、PQBP5の研究が進むことで、がんや老化の制御につながる可能性もある。
発表論文
Jin, X., Tanaka, H., Jin, M., Fujita, K., Homma, H., Inotsume, M., Yong, H., Umeda, K., Kodera, N., Ando, T. & Okazawa, H. (2023)
PQBP5/NOL10 maintains and anchors the nucleolus under physiological and osmotic stress conditions.
Nat Commun 2023, 14 (1), 9. doi: 10.1038/s41467-022-35602-w
PQBP5/NOL10 maintains and anchors the nucleolus under physiological and osmotic stress conditions.
Nat Commun 2023, 14 (1), 9. doi: 10.1038/s41467-022-35602-w