
脊髄小脳変性症7型遺伝子Ataxin-7の新たな機能の発見 (2011)
私たちの研究室は、アルツハイマー病、パーキンソン病に次いで頻度の高い神経変性疾患であるポリグルタミン病の病態解明に取り組んでいる。異常タンパクの産生、凝集体形成、神経細胞機能障害、神経細胞死に至る過程は基本的に全ての変性疾患において共有している。
脊髄小脳変性症7型は小脳失調に加えて網膜色素変性を伴う点で特徴的な常染色体優性遺伝の変性疾患である。その原因遺伝子であるAtaxin-7は酵母タンパクSgf73のホモログと考えられている。Sgf73はクロマチンリモデリング複合体であるSAGA複合体の構成要素である。SAGA複合体にはSpt, AdaおよびGcn5 acetylaseが含まれており、ヒストンタンパク質のアセチル化を行う。このhistone acetyltransferase (HAT)活性によって、転写、DNA損傷修復、DNA複製などに多面的に関与している。哺乳類におけるSAGA複合体のホモログはTFTC, STAGA, PCAF/GCN5の3種類が知られており、Ataxin-7はTFTC複合体とSTAGA複合体に含まれることが質量解析から明らかになっている。さらに、Ataxin-7の機能低下が、網膜形成・維持に必要な遺伝子群の発現低下につながることも示されている。
このように、核におけるAtaxin-7の機能が明らかになる一方で、Ataxin-7が細胞質にも存在することが報告されている。それは死後脳あるいは培養細胞における結果であり、後者においてはNLSあるいはNES配列が、細胞質局在と関連していることも報告されている。しかしながら、細胞質に存在するAtaxin-7が機能的意味を持つかどうかは明らかではなかった。
脊髄小脳変性症7型は小脳失調に加えて網膜色素変性を伴う点で特徴的な常染色体優性遺伝の変性疾患である。その原因遺伝子であるAtaxin-7は酵母タンパクSgf73のホモログと考えられている。Sgf73はクロマチンリモデリング複合体であるSAGA複合体の構成要素である。SAGA複合体にはSpt, AdaおよびGcn5 acetylaseが含まれており、ヒストンタンパク質のアセチル化を行う。このhistone acetyltransferase (HAT)活性によって、転写、DNA損傷修復、DNA複製などに多面的に関与している。哺乳類におけるSAGA複合体のホモログはTFTC, STAGA, PCAF/GCN5の3種類が知られており、Ataxin-7はTFTC複合体とSTAGA複合体に含まれることが質量解析から明らかになっている。さらに、Ataxin-7の機能低下が、網膜形成・維持に必要な遺伝子群の発現低下につながることも示されている。
このように、核におけるAtaxin-7の機能が明らかになる一方で、Ataxin-7が細胞質にも存在することが報告されている。それは死後脳あるいは培養細胞における結果であり、後者においてはNLSあるいはNES配列が、細胞質局在と関連していることも報告されている。しかしながら、細胞質に存在するAtaxin-7が機能的意味を持つかどうかは明らかではなかった。
私たちは今回、Ataxin-7とDsRedなどの蛍光タンパクの融合タンパク質の発現を通じて、Ataxin-7が微小管と共局在すること、Ataxin-7が微小管の安定化に寄与していることを示した。
まず、正常な状態でAtaxin-7を発現することが知られているHela細胞をAtaxin-7抗体で染色すると、細胞周期に関連して、M期の細胞の細胞質に内在性Ataxin-7の線維状の染色像が認められた。次に、Ataxin-7-DsRed融合タンパク質を培養細胞に発現させてlive imagingで観察すると、Ataxin-7-DsRedが核と細胞質の局在をダイナミックに変化させていることを認めた。Ataxin-7-Mycの局在は免疫染色でalpha-tubulinと重なっており(図1)、免疫沈降においても両者がassociateしていることが示された。 alpha-tubulinとの結合に正常型Ataxin-7と変異Ataxin-7の差はなかった。さらに、nocodazoleで処理すると、Ataxin-7が正常型と異常型の差異なく微小管を安定化することが認められ、siRNAのAtaxin-7ノックダウンで安定性がより低下することと合致していた。また、異常Ataxin-7トランスジェニックマウスを用いてAtaxin-7を染色すると、正常マウスに認められるプルキンエ細胞樹状突起のAtaxin-7が消失して、Ataxin-7凝集体内に取り込まれている像が認められた。これらのことから、封入体へのAtaxin-7の取り込みが、微小管の不安定性につながっている可能性が示唆された(下図)。
まず、正常な状態でAtaxin-7を発現することが知られているHela細胞をAtaxin-7抗体で染色すると、細胞周期に関連して、M期の細胞の細胞質に内在性Ataxin-7の線維状の染色像が認められた。次に、Ataxin-7-DsRed融合タンパク質を培養細胞に発現させてlive imagingで観察すると、Ataxin-7-DsRedが核と細胞質の局在をダイナミックに変化させていることを認めた。Ataxin-7-Mycの局在は免疫染色でalpha-tubulinと重なっており(図1)、免疫沈降においても両者がassociateしていることが示された。 alpha-tubulinとの結合に正常型Ataxin-7と変異Ataxin-7の差はなかった。さらに、nocodazoleで処理すると、Ataxin-7が正常型と異常型の差異なく微小管を安定化することが認められ、siRNAのAtaxin-7ノックダウンで安定性がより低下することと合致していた。また、異常Ataxin-7トランスジェニックマウスを用いてAtaxin-7を染色すると、正常マウスに認められるプルキンエ細胞樹状突起のAtaxin-7が消失して、Ataxin-7凝集体内に取り込まれている像が認められた。これらのことから、封入体へのAtaxin-7の取り込みが、微小管の不安定性につながっている可能性が示唆された(下図)。
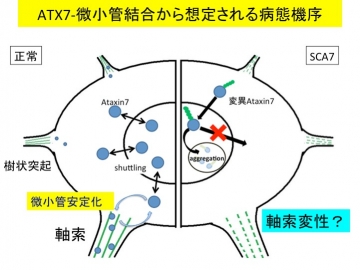
Ataxin-7が凝集して細胞質に出れなくなるとtubulinとの結合を介した微小管安定化が出来なくなる。
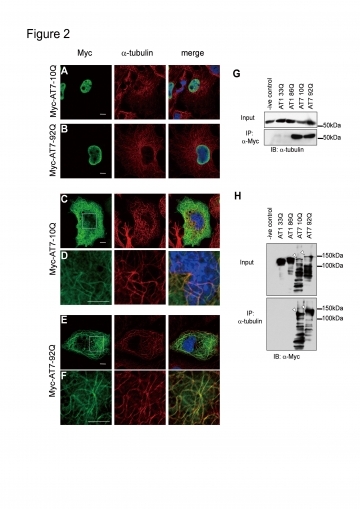
正常型および変異Ataxin-7はalpha-tubulinと結合する。
発表論文
Nakamura, Y., Tagawa, K., Oka, T., Sasabe, T., Ito, H., Shiwaku, H., La Spada, A.R. and Okazawa, H. (2012).
Ataxin-7 associates with microtubules and stabilizes the cytoskeletal network.
Hum Mol Genet. 21 (5): 1099-1110. doi: 10.1093/hmg/ddr539
Ataxin-7 associates with microtubules and stabilizes the cytoskeletal network.
Hum Mol Genet. 21 (5): 1099-1110. doi: 10.1093/hmg/ddr539