
研究概要
胚発生とは、ひとつの受精卵が細胞分裂を繰り返しながら多種・多様の細胞に分化していき、機能的な複合体を構成していくなかで最終的に完成された生物個体になる過程をさす。初期発生において均一な細胞集団は、ある特定の時期に特定の領域の細胞が特殊化して、たとえば頭部を形成したり、腹や背中を形成したりする。こうした細胞の運命決定は、個々の細胞内に備わるシグナル伝達経路が細胞外に存在する様々なシグナルにより活性化され、それに適した応答をすることにより実施される。
また、胚発生の段階で機能する細胞内情報伝達経路は成体においても様々な機能を果たしており、癌化を含めた疾患の要因となることが知られている。このことは、発生過程におけるシグナル伝達経路の理解を深めることを通して得られる知見が種々の疾患の発症機構を明らかにすることへつながるものと考えられる。
我々は、発生過程における細胞の運命決定において重要な役割を担っている細胞外シグナル分子、Wntファミリーに注目し、これらの因子が引き起こす細胞内シグナル伝達経路の解析を行っている。また、これらの研究を進める過程で同定した偽性低アルドステロン症2型原因遺伝子WNKにも注目し、その機能解析を進めている。これらの研究において、分子生物学的、生化学的解析に加え、実験材料として胚発生のモデル動物であるアフリカツメガエル(Xenopus laevis)を用いた機能解析を行っている。
また、胚発生の段階で機能する細胞内情報伝達経路は成体においても様々な機能を果たしており、癌化を含めた疾患の要因となることが知られている。このことは、発生過程におけるシグナル伝達経路の理解を深めることを通して得られる知見が種々の疾患の発症機構を明らかにすることへつながるものと考えられる。
我々は、発生過程における細胞の運命決定において重要な役割を担っている細胞外シグナル分子、Wntファミリーに注目し、これらの因子が引き起こす細胞内シグナル伝達経路の解析を行っている。また、これらの研究を進める過程で同定した偽性低アルドステロン症2型原因遺伝子WNKにも注目し、その機能解析を進めている。これらの研究において、分子生物学的、生化学的解析に加え、実験材料として胚発生のモデル動物であるアフリカツメガエル(Xenopus laevis)を用いた機能解析を行っている。
Wntシグナル伝達経路の解析
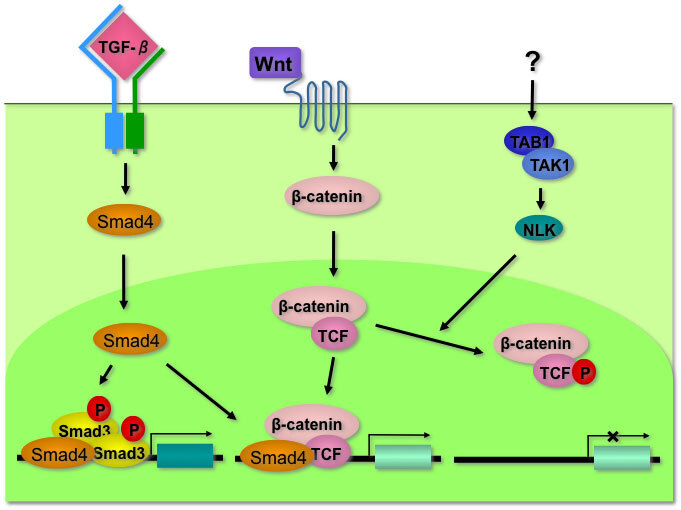
Wntシグナル伝達経路は種々の生物において高度に保存されたシグナル伝達経路であり、ガンや胚発生において重要な役割を担っている。Wntシグナル伝達経路はその作用機序から (1)β-catenin/TCFを介した転写活性化経路 (canonical)、(2)カルシウム流入を介したシグナル経路 (non-canonical)、(3)Rho、JNKを介し、細胞極性に関わるPCP経路(non-canonical)の3つに分類される。canonical Wntシグナル伝達経路は、リガンドであるWntと膜タンパク質であるFrizzled及びLRPとの結合により始まる。Wntリガンド非存在下では、APC/Axin/GSK-3β複合体のGSK-3βによりβ-cateninはリン酸化、ユビキチン化され、プロテアソームにおいて分解される。Wntリガンド存在下ではDVLが細胞膜で活性化され、APC/Axin/GSK-3β複合体を不活化する。分解されなかった細胞質中のβ-cateninは核内へ移行し、転写因子Tcfと結合し、Wntの下流(標的)遺伝子の転写を活性化する。
これまでにcanonical Wntシグナル伝達経路に関わる分子として、NLK(Nature 1999), Smad4(Nature 2000), IQGAP1(J.B.C. 2013)等を同定し、その機能を明らかにしてきた。
偽性低アルドステロン症II型原因遺伝子WNK
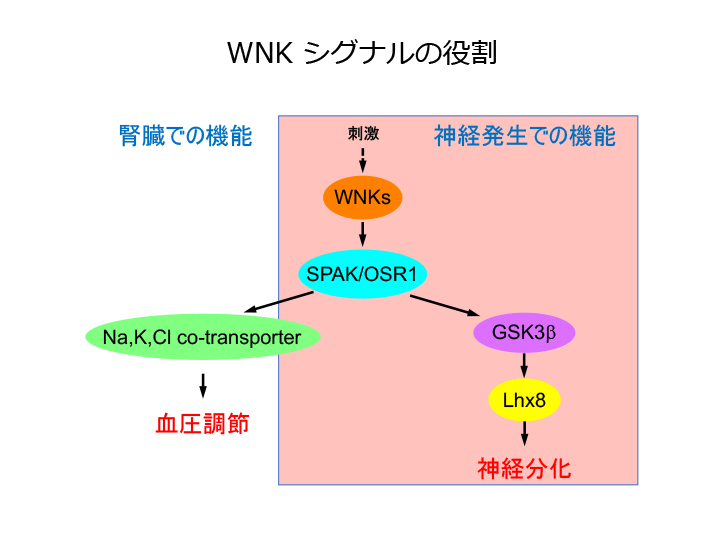
偽性低アルドステロン症II型(PHAII)は高カリウム血症と高血圧を呈する常染色体優性遺伝性疾患であり、原因遺伝子としてプロテインキナーゼWNK1およびWNK4が同定された。我々は、WNK1結合因子としてSTE20様プロテインキナーゼSPAK/OSR1を同定し、機能解析を進め、生体内においてWNK1→SPAK/OSR1→共輸送体というシグナル伝達経路が存在し、この経路の制御異常が高血圧発症の一因である可能性を示した(J. Biol. Chem. 2005, Cell Met. 2007)。
WNKシグナルによる神経分化への関与
一方、WNK1遺伝子は遺伝性感覚性自律神経性ニューロパチー(HSAN)II型の原因遺伝子でもあることが示され、我々はショウジョウバエのWNK相同遺伝子欠損変異体を樹立し、遺伝学的解析等を進め、WNKがGSK3分子を経てLhx8遺伝子の発現を介することで神経分化に関わることを明らかにした(PLos One 2013, 2018)。
WNKによるβ-cateninの分解制御
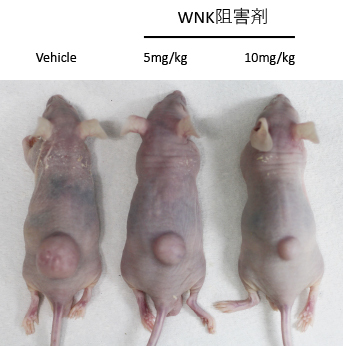
我々は以前WNKをGSK3β結合因子として単離したことから、Wntシグナルに対するWNKの関係を調べた結果、WNKがE3リガーゼであるMAEAとRMND5Aを介したユビキチン化によるβ-cateninの分解制御に関わることを明らかにした。さらに、大腸がんで観察される活性化型のβ-catenin変異体(通常のリン酸化、ユビキチン化による分解制御を逸脱したもの)も、WNKをノックダウンすることで分解されることから、WNKの発現をノックダウン、もしくは機能を阻害することができれば、大腸がんでのWntシグナルの異常な活性化を抑制でき、抗腫瘍効果を発揮することが期待された。そこで、これまで血圧調節を目的として報告されてきたWNK阻害剤を用い、大腸がん細胞を移植したマウスに投与し抗腫瘍効果を調べたところ、ある1種のWNK阻害剤を投与したマウスでは、濃度依存的に大腸がんの大きさが縮小すること、実際にそれらの腫瘍ではβ-cateninの量が抑制されていることを明らかにしました(Com. Biol. 2020)。