
発生段階依存的選択的スプライシング
Ohno et al., Genes & Development (2008)
線虫のコラーゲン遺伝子の発生段階依存的選択的スプライシングの制御機構について紹介します.
線虫のコラーゲン遺伝子の発生段階依存的選択的スプライシングの制御機構について紹介します.
この研究成果は、Gene & Development誌(2008年22巻3号)の表紙となり、巻頭の展望記事で詳しく紹介されました.
この研究成果は、東京医科歯科大学からプレスリリースされ、毎日新聞2008年1月30日付朝刊で報道されました.
この研究成果は、Nature Reviews Genetics誌およびNature Reviews Molecular Cell Biology誌の2008年3月号でそれぞれ「Colour-Changing Worms」として報道されました.
この研究成果は、東京医科歯科大学からプレスリリースされ、毎日新聞2008年1月30日付朝刊で報道されました.
この研究成果は、Nature Reviews Genetics誌およびNature Reviews Molecular Cell Biology誌の2008年3月号でそれぞれ「Colour-Changing Worms」として報道されました.
線虫基底膜コラーゲン遺伝子
線虫let-2遺伝子は基底膜を構成するコラーゲンタイプIVのアルファ2鎖をコードし、エクソン9とエクソン10が相互排他的で、発生段階依存的に制御されます.胚期には9型のみが発現し、発生が進むに連れて徐々に切り替わり、成虫期は10型のみが発現しています.
let-2スプライシングレポーター
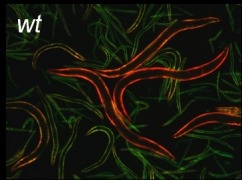
let-2レポーター線虫
図のようなレポーターミニ遺伝子を作製し、内在性のlet-2遺伝子が主に発現する体壁筋に発現させたところ、胚では9型(GFP)のみを発現し緑色をしていますが、発生段階依存的に徐々に色が変わり、成虫では10型(RFP)のみを発現して赤くなるレポーター線虫が得られました.
変異体の単離と制御因子の同定
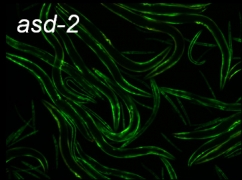
asd-2変異体
変異体スクリーニングの結果、成虫になっても胚型(GFP)が優勢に発現し緑色である変異体株を単離しました.
そして、原因遺伝子のマッピングを行い、新規のSTARファミリーRNA結合タンパク質が発生段階依存性の制御因子であることを見出して、ASD-2(Alternative-Splicing- Defective-2)と命名しました.
そして、原因遺伝子のマッピングを行い、新規のSTARファミリーRNA結合タンパク質が発生段階依存性の制御因子であることを見出して、ASD-2(Alternative-Splicing- Defective-2)と命名しました.
シスエレメントの同定
ミニ遺伝子の改変を利用して、イントロン10にあるCUAACの繰り返し配列が胚型から成虫型への発生段階依存的な切り替えに必要なシスエレメントであることを見出しました.
さらに、ASD-2がこのCUAACの繰り返し配列に特異的に結合することを確認しました.
さらに、ASD-2がこのCUAACの繰り返し配列に特異的に結合することを確認しました.
プロセシング中間体の同定
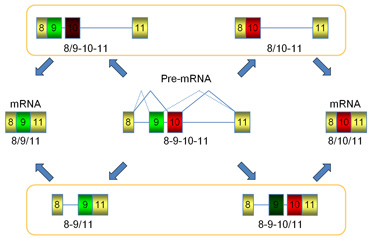
let-2 mRNA前駆体の仮想的プロセシング経路
let-2遺伝子のエクソン8からエクソン11に着目すると、mRNA前駆体からエクソン9型または10型の成熟mRNAが生成されるには、図のように、選択的エクソンの上流のイントロンと下流のイントロンが除かれる合計2度のスプライシングが必要となります.
そこで、理論上4種類存在する、1つのイントロンのみが除去された「プロセシング中間体」の量を、放射性同位元素を利用したRT-PCRにより定量し、野生型とasd-2変異体で比較しました.
その結果、胚期ではエクソン9からエクソン11へスプライシングした中間体のみが検出されること、野生型では成虫期にエクソン10からエクソン11へスプライシングした中間体に切り替わること、asd-2変異体では、成虫期でも、胚型が主要な中間体であることがわかりました.
そこで、理論上4種類存在する、1つのイントロンのみが除去された「プロセシング中間体」の量を、放射性同位元素を利用したRT-PCRにより定量し、野生型とasd-2変異体で比較しました.
その結果、胚期ではエクソン9からエクソン11へスプライシングした中間体のみが検出されること、野生型では成虫期にエクソン10からエクソン11へスプライシングした中間体に切り替わること、asd-2変異体では、成虫期でも、胚型が主要な中間体であることがわかりました.
制御のモデルと研究成果の意義
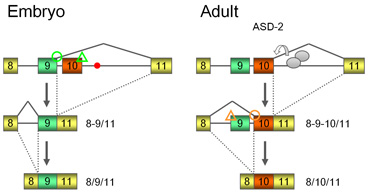
let-2遺伝子の制御のモデル(Genes Dev, 2008より)
let-2の相互排他的選択的スプライシングでは、選択的エクソンの下流のイントロンが先行して除去されます.すなわち、胚ではエクソン9とエクソン11の間でまずスプライシングが起こり(図左)、成虫期では、ASD-2のはたらきによりイントロン10の除去が促進されます(図右).
この研究は、スプライシング部位の相対的な強弱や制御因子によるスプライシングの促進機構が連動してイントロン除去の順序を制御することによりスプライシング・パターンの厳密な切り替えが実現していることを、遺伝学的な実験系で、内在性の遺伝子を用いて初めて明確に示したもので、mRNAプロセシングの運命決定過程の解析に新たな道を拓くものです.
この研究は、スプライシング部位の相対的な強弱や制御因子によるスプライシングの促進機構が連動してイントロン除去の順序を制御することによりスプライシング・パターンの厳密な切り替えが実現していることを、遺伝学的な実験系で、内在性の遺伝子を用いて初めて明確に示したもので、mRNAプロセシングの運命決定過程の解析に新たな道を拓くものです.