石野研究室
石野研究室
東京医科歯科大学難治疾患研究所・エピジェネティクス分野
〒113-8510 東京都文京区湯島1-5-45tel: 03-5803-4862 fax: 03-5803-4863
(医歯学総合研究棟II期 23階 map)
石野研究室1984年 Surani らによって、人工的に単為発生胚が作製され、それらは胎盤形成不全によって初期胚致死になることが示された。単為発生胚が初期胚致死となる原因は、父性発現インプリンティング遺伝子の発現欠如によると考えられる。そこで、新規父性発現インプリンティング遺伝子のスクリーニングを行ない、Peg10 ( Paternally expressed gene 10 ) の単離に成功した。非常に興味深いことに、 Peg10 は、 ORF1 と -1 フレームシフトを介して翻訳される ORF2 の2つのタンパク質をコードし、それぞれがフグの sushi-ichi レトロトランスポゾンの GAG、 POL タンパク質に高い相同性を持つレトロトランスポゾン由来の遺伝子であった。しかしながら、 Peg10 は他のレトロトランスポゾンのように不活性化されずに胎盤で高発現し、哺乳類で高度に保存されていることから、発生初期の胎盤形成に必須な遺伝子、すなわち、単為発生胚が初期胚致死になる原因遺伝子であると考えた。そこで、私たちは、 Peg10 KO マウスの作製により、実際に Peg10 が発生初期に必須かつ単為発生胚が初期胚致死になる原因遺伝子であることを証明することを試みた。
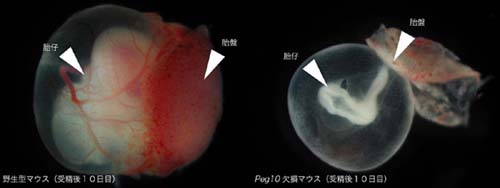
驚くべきことに、 Peg10 (+/-) マウスは、受精後 10 日目までに、初期胚致死の表現型を示した。 Peg10 は、父性発現インプリンティング遺伝子であることから、父親由来で KO アレルの伝わった Peg10 (+/-) マウスは初期胚致死になるが、母親由来で KO アレルの伝わった Peg10 (-/+) マウスは正常に生まれてくる。雄の Peg10 (-/+) マウスと交配した雌を妊娠初期で帝王切開すると、 Peg10 (+/-) マウスには胎盤が全く形成されずに致死となっていることがわかった。これは、レトロトランスポゾン由来の遺伝子である Peg10 が哺乳類の単為発生胚が初期胚致死となる原因になっていることを意味し、さらに、レトロトランスポゾンが哺乳類の進化の過程で個体発生に必須な機能を獲得し、内在性の遺伝子として働いていることを示す初めての報告である。このことから、哺乳類の胎生という個体発生様式に必須な臓器である胎盤が、進化の過程で獲得されるために、レトロトランスポゾンに由来した遺伝子が使われていることを意味している。 すなわち、Peg10は現在の哺乳類の個体発生に必須な遺伝子であり、哺乳類の進化を考える上でも重要な意味を持つ遺伝子であることが示された。