 Gregor Mendel (1822-1884) 肖像は1862年
Gregor Mendel (1822-1884) 肖像は1862年1.メンデルの遺伝の法則 2.メンデルの法則が当てはまらない場合 3.遺伝学の応用 関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください) 遺伝学電子博物館(国立遺伝学研究所作成) 更新日:2005/05/09
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
メンデルの法則の概要
メンデルの著した「植物雑種の研究」原典(ドイツ語版、英語版、英語注釈つき)
メンデルは1857年からエンドウを注意深く何代も観察して、親の特徴が雑種の子孫に再び表れるときに、あるパターンがあることに気がついた。この法則性を明らかにしようとして、実験を行うことにした。
Gregor Mendel (1822-1884) 肖像は1862年
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
メンデルの生涯
メンデル以前に、遺伝の概念がなかったわけではもちろんない。ただ交配を行っても、明瞭な法則性を見出すことには、誰も成功していなかった。その理由は、純系を使った交配実験が行われたのではなく、最初から雑種を使っていたので、結果が明瞭ではなかったのである。
彼は、実際の実験を始める前に数多くの遺伝的な特徴について、純系を得るための作業をおこなった。そして最終的に7つの、明らかに対照的な(対立する)特徴(形質、caharacter)を持つ種子を選んだ。
1)黄色と緑色の種子(seed
color)、
2)丸い種子としわあるの種子(seed shape)、
3)黄色と緑色のさや(pod color)、
4)背丈が高いか低いか(stem length)、
5)さやが膨らんでいるか平たいか(pod shape)、
6)花の色が紫色か白色か(flower color)、
7)花が茎の頂端につくか茎全体につくか(flower position
on stem)、
である。
1)優劣の法則
まずはじめに、対立形質を持つ2つの純系の植物を交雑する実験をおこなった。
たとえば、紫色の花を咲かせる植物と白い花を咲かせる植物(これをP世代という、parental generation)を交雑させると、全ての子(これをF1世代という、first filial generation)は紫色の花を咲かせた。
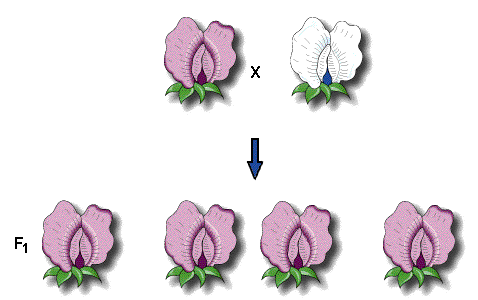
花の色という形質のうち、「紫色」「白色」を区別するときは、それぞれをtraitという。日本語では形質といって区別しないことが多い。
F2世代(second filial generation)は、F1世代の交雑や、自家受粉によって作ることができる。その結果、F2世代では、705の紫色の花を咲かせる植物と、224の白色の花を咲かせる植物が得られた。白色の花を咲かせる遺伝因子はF2世代に再び出現するので、F1世代で失われたのではなかったのだ。
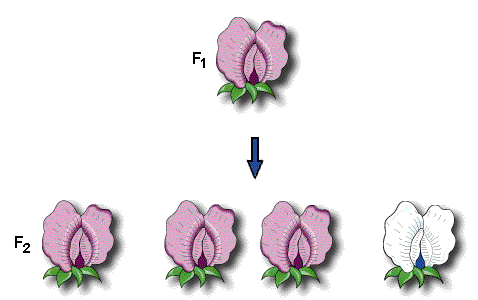
紫色の花を咲かせる植物と白色の花を咲かせる植物の出現頻度は705:224=3.15:1で、整数にするとほぼ3:1となる。このようにメンデルは常に標本の数を多数取り、それを数量的にあつかった。
その他の形質についても同様な実験を行い、いずれの場合もF1では一方の形質のみが現れ、次のF2ではほぼ3:1になることがわかった。
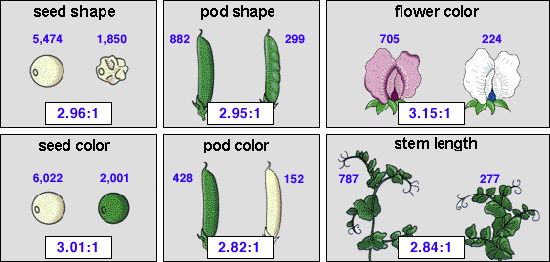
この実験から、今では優劣の法則、分離の法則として知られる2つの法則が見出され、なぜその様なことが起こるのかについて、合理的な説明がなされた。
メンデルは、形質を支配する独立した要素があると考えた。もちろんメンデルは、遺伝子という言葉は使っていない。遺伝的な要素という言葉を使っている。しかし、これは現在の遺伝子とほとんど同義なので、ここでは遺伝子という用語を使うことにする。遺伝子は、基本的には一対、存在し、それぞれ父親と母親から一つずつ受け継ぐと考える。
メンデルは、F1では片親からの遺伝子が、もう一方の親の遺伝子の性質を覆い隠してしまうと考えた。上の例では、「紫色の花の色」という性質が優性(dominant)で子に現れ、「白色の花の色」という性質が劣性(recessive)で隠されてしまうのである。現在、優劣の法則として知られている概念である。
なお、ここでいう優性と劣性は、性質が優れている劣っているということとは関係ない。英語にあるように現れると隠れるという意味である。そのため顕性と潜性と言う言葉が使われることもあるが、ここでは優性劣性を使うことにする。
2)分離の法則
メンデルは、配偶子が形成される時に、2つの遺伝子は素粒子のようにふるまい、生殖細胞には各々の対の片方しか含まないように分かれるのだと主張した。この過程の間、2つの遺伝子は、なんの変更も受けず、混じり合うこともない。遺伝子は消えたり混じったりしないので、劣性の性質は覆い隠されるだけで、次の世代にふたたび表れることが可能なのである。この考え方が対立遺伝子の分離の法則である。
ここではエンドウの代わりに、モルモットの毛色の遺伝を例にとってみよう。純系の黒色雌モルモットと、茶色の雄モルモットを交雑してできた子供はすべて黒色である。つまり、茶色は劣性であって、黒色は優性遺伝子なのである。ここで、黒遺伝子を優性B、茶色遺伝子を劣性bとあらわす。個体の形質は表現型(phenotype)といい、遺伝的構成は遺伝子型(genotype)という。つまり、母親の遺伝子型は優性ホモBBであり、父親の遺伝子型は劣性ホモbbであったのである。
黒色雌のモルモットから生じる卵細胞はいずれも遺伝子Bをもつ。一方、茶色雄モルモットから生じる精子細胞は遺伝子bをもつ。そのため受精によってできる子供はすべて、遺伝子型がBbで表現型が黒色のモルモットとなる。
この遺伝子型の雌がつくる卵の遺伝子はBとbは1:1、同じく雄がつくる精子もBとbが1:1である。これらは同数だけ作られ、同形の対立遺伝子を持つ精子や卵の間で、特別な誘引や反発作用はおこらないので、受精はランダムに起こる。
したがってこれらの卵と精子が受精すれば次のような遺伝子型を持った子供が生まれる。
雑種第一代によってつくられる生殖細胞 |
卵(B) |
卵(b) |
精子(B) |
BB |
Bb |
精子(b) |
Bb |
bb |
F2の子供のうち4分の3は遺伝子の型としてBB型もしくはBb型で、外見的には黒である。残り4分の1が、遺伝子型がbbで外見的に茶色である。これが、メンデルのエンドウの交配実験によって得られた、約3:1というF2の比率の遺伝的メカニズムなのである。
表現系では黒:茶=3:1であるが、遺伝子型の比率はBB型:Bb型:bb型=1:2:1である。
現在では、メンデルが仮定した素粒子のような要素は、実在する染色体上の遺伝子座として認識されている。分離の法則で分かれると書いたのは、減数分裂のときに染色体がおのおのの生殖細胞に分配されることに対応している。
3)独立の法則
単純な一遺伝子雑種(monohybrid cross)は、それぞれ、ある一つの座を示す対立遺伝子の一対が関係する現象である。メンデルは、二つ(dihybrid cross)またはそれ以上の形質をもつ交配種についても分析した。二つの対立遺伝子が異なった染色体の上にある時、それぞれの対はそれぞれ独立して分配されるのである。
ふたたびモルモットを例に取ろう。優性ホモ(同形接合)の黒色で毛の短いモルモット(BBSS型)と劣性ホモの茶色で毛の長いモルモット(bbss型)が交配された時、BBSS型の動物はすべてBS型の生殖細胞を生じ、bbss型の個体はすべてbs型の生殖細胞を生じる。
これらの子供はすべてBbSsの遺伝子型を持つので、毛色および毛の長さについて異形接合体であり、みな黒く短い毛をしている。
F1個体は4種類の配偶子を等しい確立で作り出す(BS、Bs、bS、bs)。そのため、これらの配偶子の交雑の結果、9:3:3:1の確率で、それぞれ黒色で短い毛、黒色で長い毛、茶色で短い毛、茶色で長い毛というF2世代が生まれる。
雑種第一代による生殖細胞 |
卵(BS) |
卵(Bs) |
卵(bS) |
卵(bs) |
精子(BS) |
BBSS |
BBSs |
BbSS |
BbSs |
精子(Bs) |
BBSs |
BBss |
BbSs |
Bbss |
精子(bS) |
BbSS |
BbSs |
bbSS |
bbSs |
精子(bs) |
BbSs |
Bbss |
bbSs |
bbss |
この結果を踏まえて、メンデルは今では独立の法則として知られる遺伝法則を公式化した。各々の配偶子は、各々の座の一つの対立遺伝子を含み、異なる座の対立遺伝子とは互いに干渉することなく、ランダムに配偶子に分配される。上の例で言えば、B(あるいはb)とS(あるいはs)はお互いに独立して、配偶子に分配されると言うことである。
メンデルの遺伝の法則は、発表された当時(1865)は誰からも理解されなかった。早すぎた天才の悲劇である。1900年になってド・フリース、チェルマク、コレンスの3人によって、独立にメンデルの法則が再発見され、コレンスによって3つの法則にまとめられた。その後、染色体研究の進展に伴い、遺伝子が染色体上の実体であることが認められ、現在の遺伝子の考えに発展する。メンデルはその一番初めの基礎を築いた人である。
4)ヒトに見られるメンデル遺伝
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
http://www.sonic.net/~nbs/projects/bio115l/form.html
その後、植物だけでなくあらゆる生物で、遺伝の法則が成り立つことが明らかになっていった。次の節で述べるように、メンデルの遺伝の法則に単純に当てはまるものは少ないが、ヒトでも上に述べた法則に従う形質がある。
ヒトの場合は、交配実験を行うことはできないので、家系分析をして推定する。たとえば、髪の毛の生え際が富士額(widow's speak)かそうでないかは、メンデル遺伝に従う。富士額は優性でWで表し、富士額を示さずストレートのほうはwで表す。
一方、耳たぶの形で、耳たぶがたれ下がっているほうが優性Fで、耳たぶがくっついているほうが劣性fである。
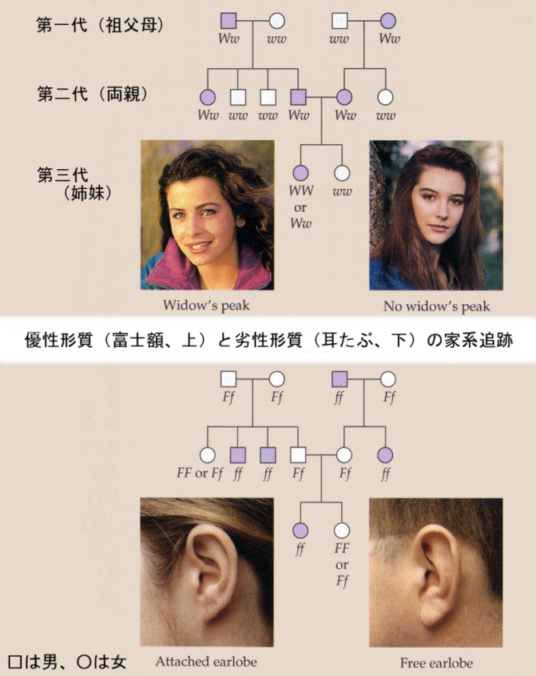
この他、舌を巻いてU字型にできるか(tongue rolling)、親指を立てたとき親指を外側にそらせられる角度(hitchhiker's thumb)が大きいか、指を交互にして両手を組んだとき左手の親指が上になる(interlocking fingers)か、などがある。いずれも記述したほうが優性で、逆が劣性である。
クラスで調べて頻度を取ったり、両親と自分で各形質のtraitsを家系追跡をして、自分がどのような遺伝子をもつか調べてみよう。
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
人類遺伝学講義(ロンドン大学)
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
http://www.biology.iupui.edu/biocourses/N100H/ch10genetics.html
メンデルが選んだ7つの形質は、はっきりとした対立形質だったというだけでない。独立の法則があてはまるような形質が選ばれている。おそらくメンデルは慎重に7つの形質を選び出したのであろう。実際に、エンドウの染色体数は2n=14で、1つの例外を除いてそれぞれの形質は、別の染色体上にあることが、後で明らかになった。
遺伝子が、はっきりとした対立形質であり、かつ別の染色体上にあることが、メンデルの遺伝の法則が成り立つためには必要である。そのために、その後、例外がたくさんあることが明らかになる。現在では、一般的な形で成り立つのは、分離の法則だけで、それ以外は法則とは言えないと考える人もある。
しかしながらこのことは、メンデルの考えが誤っていたことを意味するのではない。むしろメンデルの実験家としての資質が優れていたことを示している。これらの例外は、いずれも染色体の構造や細胞分裂のときの挙動、遺伝子本体のDNAとタンパク質の関係が明らかになって、素直に理解できるものである。
補足:遺伝子座の命名法
ここで、遺伝子の表現方法について補足しておく。メンデルは遺伝子を表すのに、アルファベットの一文字を使い、大文字を優性、小文字を劣性とした。その後、モーガンたちは突然変異によって生じた遺伝子座の名前を、その特徴を表すアルファベット1-3文字で表し、その対立形質の野生型(wild
type)をその文字列に+を上付きで付する方法を考案した。突然変異が劣性であれば小文字で、優性であれば最初の文字を大文字で示す。
ショウジョウバエを使って、突然変異の研究をしていたモーガンらが見つけた、劣性突然変異体に翅が痕跡的なものがあった。これは劣性だったので、痕跡的な(vestigial)翅と言うので、遺伝子座はvgと名づけられた。したがって野生型はvg+である。
遺伝子座は相同染色体上に一対あるので、優性ホモの野生型はvg+vg+と表し、痕跡翅を持つ劣性ホモの個体はvgvgと表す。区切りがわかりにくいのでスラッシュを入れる場合もある。その場合はそれぞれ、vg+/vg+とvg/vgと表す。さらに遺伝子座の対応が明らかなときには、上付きとせずに+だけで表すこともある。この場合は、+/+とvg/vgとなる。
1)連鎖(linkage)
まず独立の法則が当てはまらない例として、連鎖を取り上げよう。
すでに述べたように、遺伝子は染色体上にあり、しかも1本の染色体上には、複数の遺伝子が存在する。染色体は減数分裂のときに1つの単位として行動するので、同じ染色体上に載っている全ての遺伝子は、まとまって受け継がれる傾向にある。
もし、染色体のひとまとまりが決して変化しないならば、染色体上の遺伝子も常にまとまって受け継がれるであろう。したがって同じ染色体上にある遺伝子では、メンデルの独立の法則は当然、当てはまらない。
さて、ショウジョウバエにおいて、翅の形(正常翅は優性遺伝子であるvg+で、痕跡翅は劣性遺伝子であるvgで表す)を支配する座と、体の色(灰色はb+で、黒色はbで表す)を支配する座は、同一染色体上に位置している。そのため、両者はいっしょになって受け継がれる傾向がある。これを、連鎖(linkage)していると言う。
優性ホモのb+b+vg+vg+バエ(野生型)と劣性ホモのbbvgvgバエを交雑すると、F1ではこれらの遺伝子に関してヘテロ接合体のハエb+bvg+vgが生まれ、表現型は正常翅灰色である。
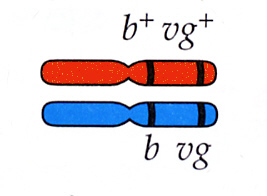
F2を考えてみよう。もし、これらの特徴を支配する座が連鎖していないならば(同じ染色体上にないならば)、減数分裂の時、対立遺伝子は独立してふりわけられ、ヘテロ接合体の親からは、4種類の配偶子(b+vg+、b+vg、bvg+、bvg)が生じ、メンデルの分離の法則にしたがって9:3:3:1の比でそれぞれの表現型のハエが生まれる。
ところが完全に連鎖している(たとえば遺伝子座が隣り合っている)場合は、配偶子は2種類(b+vg+とbvg)しかできない。したがって、一遺伝子の場合と同じようになり、F2世代は3:1で正常翅灰色と痕跡翅黒色が生まれる。
しかし、実際にはこの二つの遺伝子座は隣り合っているわけではない。そのため、第1減数分裂中期に、染色体が対になったとき(対合)、染色分体の間で交叉(chiasma)が生じて乗り換え(crossing over)がおこり、その結果、遺伝子の組み換え(recombination)がおこる。そのため、ある割合でb+vgとbvg+の配偶子が生じる。そのために9:3:3:1とはずれてくるが、両親とは違った組み合わせの個体が生じる。
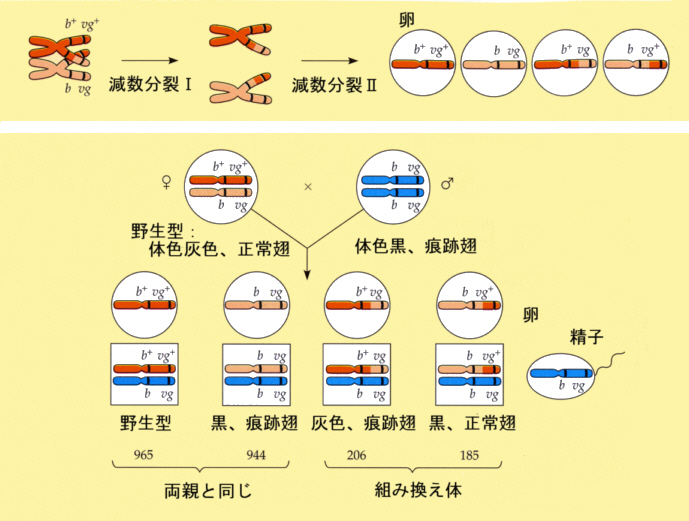
この組み換えがどのくらいの割合でおこっているかをしらべるためには、F1と劣性ホモを掛け合わせてそのこどもの表現型の比を調べると分かる。これを戻し交配(back cross)という。F1がつくる配偶子の遺伝子は、戻し交配によってb+vg+:b+vg:bvg+:bvg=965:206:185:944であることが分かった。これから組み換え率が(206+185)/(965+206+185+944)に100をかけて17%と求めることができる。
組み換え率は、連鎖している染色体上の距離に比例している。組み換え率が50%の時は連鎖していないことになる((1+1)/(1+1+1+1)より)。50%より小さければ小さいほど、二つの遺伝子の距離は近くなる。
多く形質について組み換え率を求めて、それを直線上に並べてゆくと、遺伝地図ができる。モーガンはショウジョウバエを使い、4つの連鎖群(すなわち染色体)の遺伝地図を作成した。遺伝地図(あるいは連鎖地図)は組み換え率をもとに計算した地図なので、並び方は正しいが、遺伝子座間の絶対的な距離を正確に反映しているわけではない。
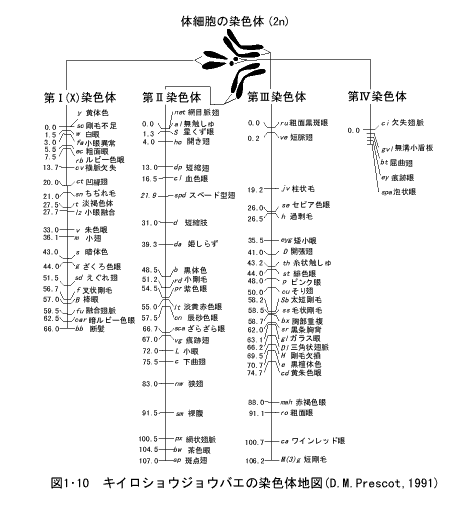
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
人の染色体地図
2)伴性遺伝(sex-linked inheritance)
性決定の仕組みはかなり様々であるが、たいていの動物は特別な性染色体を持っている。
典型的な場合、一方の性は同一な性染色体の対を持っていて、性染色体構成において、全く同一な配偶子を作り出す。他方の性は2つの異なる性染色体を持っていて、それぞれの性染色体を含む2種類の配偶子を作り出す。
ヒトも含めて多くの動物種の雌は同形配偶子性で、その細胞はX染色体と呼ばれる2つの同一の性染色体を含む。一方、雄は異形配偶子性で、その細胞はX染色体とそれよりも小さいY染色体をもっている。
正常な雄が1つのX染色体と1つのY染色体を持つ種では、これらの染色体が減数分裂の際に規則的にお互いに対合し分離する。したがってXとY染色体では、ホモ接合体の対のメンバーは大きさや形や遺伝子構成が似ているという一般規則に対する例外となる。
こうしてできた生殖細胞のうち、精子では一方がX染色体を含み、他方はY染色体を含み、全ての卵子はただ1つのX染色体を持つ。
したがって必然的に、Xを持つ卵子とXを持つ精子が受精するとXX(雌)接合体になり、卵子とYを持つ精子が受精するとXY(雄)接合体になる。
ヒトのX染色体には、両方の性に欠かせない多くの遺伝子座を含むが、Y染色体には雄に対する遺伝子を含む少数の遺伝子しか持たない。したがって、色盲や血友病のようなX染色体に位置する遺伝子によって支配される形質は、伴性遺伝をする。雌は母から1つのXを、父から1つのXを受け取る。雄は自らを雄にならしめるY染色体を父から受け取る。雄は母からただ1つのX染色体を受け継ぎ、それゆえX染色体に伴う全ての遺伝子は母から受け継いだものである。雄ではあらゆるX染色体の形質は、雌の親の遺伝子によって規定されてしまう。
3)不完全優性(incomplete dominance)
いろいろな生物の多くの形質の遺伝の研究により、対立遺伝子の対の1つの要素が、常に他方に対して完全に優性であるわけではない事がわかった。
たとえば、赤や白のオシロイバナは、日本では普通である。これらの花を交雑させると、F1の雑種はピンク色になる。このピンク色の花を交雑させると、赤、ピンク、白の花の雑種が1:2:1の割合で現れる。
ピンク色の花は明らかにヘテロ接合の個体で、赤の対立遺伝子も白の対立遺伝子も、完全には優性ではない。ヘテロ接合体がその親の中間の表現型を持つ時は、その遺伝子は不完全優性を示すと言われる。
4)複対立遺伝子
今までの例は、各々の座が最高で2つの対立遺伝子の変異型で表現され、これらの例のほとんどで、対立遺伝子の1つが優性で1つが劣性であった。
個々の二倍体の個体が特定の座に対して最大で2つの異なる対立遺伝子を持ち、半数体の配偶子が各々の座に対してただ一つの対立遺伝子を持つ例は多いが、個体群を調べると、特定の座に対して2つ以上の対立遺伝子を見い出すことがある。もし、ある座に対する3つ以上の対立遺伝子が個体群内に存在するならば、その遺伝子座は複対立遺伝子を持つと言う。
今までに示した例では、遺伝子とその表現型の関係は直接的で、明確であり、考えられている座がただ一つの形質の出現を支配していた。しかしながら形質に対する遺伝子の関係がかなり複雑な場合もある。たいていの遺伝子はおそらく多くの異なる影響、つまり多形質発現と呼ばれる性質を持つ。これは、一つの座が同時に数多くの形質に影響を与える事ができる事を示している。生物のほとんどの全ての形質が実際には多くの座によって支配されている。
遺伝子の相互作用の最も単純なタイプの一つは鶏冠の遺伝にみられる。バラ冠を表す対立遺伝子であるRは単冠の対立遺伝子であるrに対して優性である。別の遺伝子対はマメ冠を表すPと単冠をあらわすpの遺伝を支配している。それゆえ、単冠の鶏はpprrという遺伝子型をもたなけれはならない。マメ冠の鶏はPPrrかPprrのどちらかであり、バラ冠の鶏はppRRかppRrのどちらかである。ホモ接合のマメ冠の鶏とホモ接合のバラ冠の鶏を交配すると、雑種は、マメ冠でもバラ冠でもないクルミ冠と呼ばれている完全に異なるタイプになる。クルミ冠の表現型は鶏が1つか2つのR対立遺伝子と1つか2つのP対立遺伝子を持つ時にはいつでも生み出される。すなわちPPRR、PpRR、PPRr、PpRrという事である。
5)ポリジーン遺伝
背の高さ、体型、肌の色のような多くのヒトの特徴を受け継いだ構成要素は、一つの座における対立遺伝子によって受け継いだものではない。いくつかの、もしかしたら多くの異なる座の対立遺伝子が各々の特徴に影響を与える。ポリジーン遺伝という用語は多数の独立した遺伝子対が同じ特徴に対して相関的な効果を与えるときに使われる。
ポリジーンはヒトの肌の色の遺伝の原因となる。今では、4つ以上の異なる座を表す対立遺伝子が肌の色を決定するのに関わっていると考えられているが、ポリジーン遺伝の原理は3つの連鎖していない座の対立遺伝子対を用いて示すことができる(Aとa、Bとb、Cとc)。大文字は暗い肌をつくる不完全に優性な対立遺伝子を表す。大文字の数が多くなるにつれ肌がより暗くなる。
遺伝学者は望ましい表現型を示す生物を選択し、これらの生物をさらに交配する事で、商業的に好都合な特徴を持つ品種を徐々に開発している。
しかし、選択による品種の改良には限界がある。ある血統が関わっている全ての遺伝子に関してホモ接合になると、さらに選択を重ねて望ましい性質を向上させることはできなくなる。その上、近親交配の為に多数の望ましくない特徴に対しても同様にホモ接合になる事がある。
異形接合と呼ばれる完全に血のつながりのない血統間の個体を交配すると、両親よりもずっと生存に適した子孫をえることがよくある。このような改良は雑種強勢と呼ばれている。雑種強勢の一つの説明としては、各々の親の血統はある望ましくない劣性遺伝子に関してホモ接合であるが、どちらの血統も別々の望ましくない遺伝子に関してホモ接合である。各々の血統は、他方の血統の望ましくない遺伝子の埋め合わせをする優性な遺伝子を含む。そして、ある血統が、AaabbCCddという遺伝子型を持ち、別の血統がaaBBccDDという遺伝子型を持つとする。AaBbCcDdという遺伝子型を持つ雑種は、二つの親の血統の望ましい特徴の全てを発現し、望ましくない特徴は1つも発現しないのである。
![]() この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。