1.染色体 2.体細胞分裂 3.減数分裂 4.細胞周期の制御 関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください) Designer Genes http://gened.emc.maricopa.edu/bio/bio181/BIOBK/BioBookmito.html 更新日:2002/04/08
核生物の核には複数のDNA分子が存在する。それぞれのDNA分子はとても細長いので、もつれるのを防ぐために、ヒストンという筒状のタンパク質に巻き付き数珠状のヌクレオソームという構造をつくる。ヌクレオソームはさらに凝集して折り畳まれ、染色質という繊維状の構造を取っていると考えられている(細胞のプロフィール、核参照)。染色質は、60%のタンパク質、35%のデオキシリボ核酸(DNA)、5%のリボ核酸(RNA)を含む繊維状の構造である。一方、原核細胞のDNA分子は小さいので、このような構造を取らずにDNA分子のままで存在する。
細胞がDNAの情報を読み出す(転写)ときには、上に述べた折り畳み構造はほどけて、DNAが露出すると考えられている。
細胞が分裂するときには、染色質の繊維のままではまだ複雑過ぎるので、さらに凝縮して染色体という構造を取る。教科書や資料の中によく出てくるX字型をした染色体は、これからお話をする細胞分裂の中期になって明瞭になる構造であって、ふだんはあのような形をしているわけではない。DNAがどのように組織されて染色体という構造をとるか、正確なところは分かっていない。
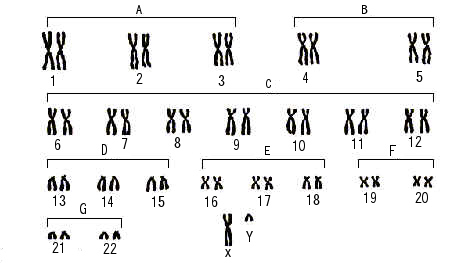
ヒト男性の染色体。女性では最後の段の性染色体がXXである。
分裂中期に見られるX字型の染色体の中央の部分をセントロメアといい、この部分で二本の染色分体(chromatid)が接している。
上の図は、セントロメアの上側を短い部分(短腕と呼びpであらわす)、下側を長い部分(長腕と呼びqであらわす)にして、大きさの順番に並べてある。こうして染色体番号を決めている。
さらに学びたい人へ(このページへ戻るときはブラウザーの戻るを選んでください)
http://arbl.cvmbs.colostate.edu/hbooks/genetics/medgen/chromo/chromosomes.html
それぞれの染色体には、非常に多くの遺伝子が座を占めている。現在のところ、遺伝子は、転写という過程でRNA分子にコピーされる領域と、転写を制御する領域をあわせたものと定義されている。遺伝子は、酵素を含むすべてのタンパク質の構造の情報をもっていて、その生合成を制御している。酵素は、触媒作用を通じて生体にとって必要な物質を合成する代謝反応を制御しているので、結局のところ、遺伝子がすべての生体の活動を制御していることになる。
ヒトゲノムプロジェクト染色体21(ヒトで一番短い染色体)の全容
http://hgp.gsc.riken.go.jp/chr21/
このように、情報の源である遺伝子を載せた染色体は、娘細胞に間違いなく分配されなければならない。真核生物の体細胞における細胞分裂には有糸分裂と呼ばれるプロセスがあり、各娘細胞にその親細胞が持つすべての染色体のコピーを確実に受け取らせている。原核生物の細胞においても、ずっと単純であるが遺伝物質の分配は正確におこなわれている。
体細胞の核には、その種に特有な数の染色体が存在する。しかしながら、種のユニークさを決めるのは、染色体の数ではなく染色体上の遺伝子のもつ情報である。上の図にあるようにヒトの場合は、常染色体22対+性染色体2本の、合計46本である。
ヒト各染色体案内
http://www.ornl.gov/hgmis/launchpad/
http://www.hgmp.mrc.ac.uk/GenomeWeb/human-gen-db-chromosomes.html
細胞はある大きさになると、成長を止めるか分裂をする。神経、骨格筋、赤血球などの細胞は、一度、成熟するとふつうは分裂しない。
細胞は、次の図のような過程で分裂する。分裂から次の分裂までを細胞周期(cell cycle)と呼び、それを一回行うのにかかる時間を世代時間と言う。この時間は、種によって異なる。細胞分裂は、核が複雑な過程をたどって分裂する有糸分裂(mitosis)と細胞質が二つに分かれる細胞質分裂(cytokinesis)にわけられる。
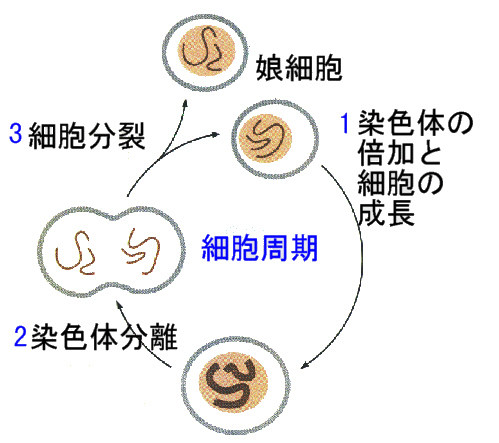
1)細胞周期
細胞周期のうち、有糸分裂の時期をM期といい、ほとんどの細胞で1時間くらい、全周期のほんの一部に過ぎない。M期とM期の間の長い期間を間期(interphase)と言う。細胞は間期の間に必要な物質を合成し、成長しており、ほとんどのタンパク質やほかの物質は間期の間ずっと合成され続けている。
間期 → 有糸分裂期(M期) → 間期
間期はさらにG1、S、G2に分けられる。S期は核のDNAの複製がおこる時期である。M期とS期の始まりまでの期間がG1期、つまり最初のギャップ期である。活発に分裂している細胞はG1期の間にDNA合成に必要な酵素が活性化される。この活性化によって細胞はふつうS期に入っていく事ができる。分裂を続けない細胞はS期に入らず、周期の外に出てG0と呼ばれるG1と似たような状態にとどまる。S期を完了すると、細胞は、二番目のギャップ期であるG2期に入る。この時期に、分裂の準備の最終段階が始まり、タンパク質の合成が増加する。
G0期
↑
G1期 → S期 → G2期 → M期
↑←←←←←←←←←←←←←←←←↓
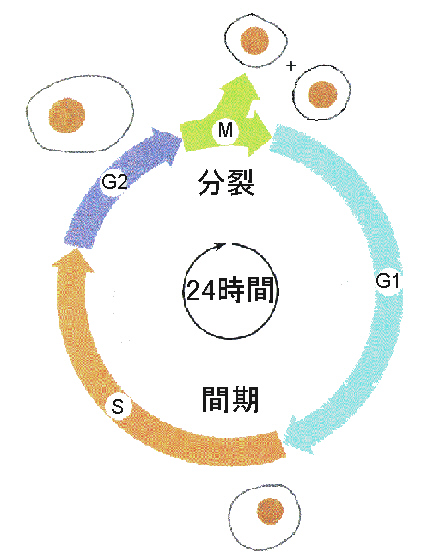
2)有糸分裂期(M期)
実際には連続した過程だが、M期は便宜的にさらに、前期(prophase)、中期(metaphase)、後期(anaphase)、終期(telophase)に分けられる。
次の顕微鏡写真は、ヌマツユクサの体細胞分裂の過程を示したものである。左から、間期、前期、中期、後期、終期を示す。この図版は広島大学教育学部理科内容学研究室が作成したもので、版権はそちらにある。同研究室が作成した植物の体細胞についてのホームページも参照するとよい。
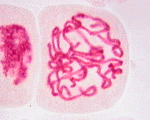

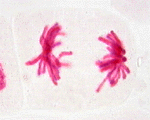
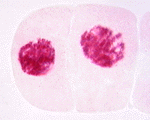
有糸分裂期の実写(WindowMediaPlayer/13M)
次に前期から細胞質分裂までの各過程を、染色体数が3本の仮想的な細胞の図(Essential Cell Biologyの図版を改変)を添えて、説明する。
●前期
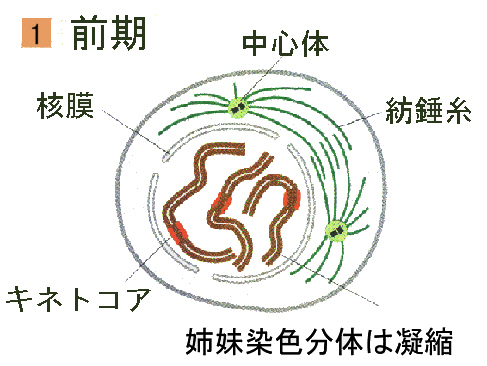
有糸分裂の最初の段階である前期になると、長い染色質の糸が凝縮し始め分裂染色体となる。この凝縮によって主に染色体が太く短くなってコイルのような状態になる。このため染色体はからみつくことなく娘細胞に分配される。それぞれの染色体はS期の間に複製されていて、姉妹染色分体と呼ばれる同一の単位の対から成っている。各染色分体には、セントロメア(centromere、動原体)と呼ばれる染まらない圧縮された領域がある。セントロメアにはキネトコア(kinetochore、こちらも動原体という語が使われる)という微小管が結合できる構造がある。
分裂している細胞は、一つの赤道面と二つの極を持つ球として表される。二つの極の間に繊維が生じ、主に微小管(microtubule)からなる複雑な構造体である紡錘体(spindle)が形成され始める。紡錘体は、後期に染色体を分離させる。それぞれの極には、微小管形成中心(MTOC)と呼ばれる、微小管が放射状に伸びていく部分がある。MTOCからはさらに微小管が多方向に伸びていって、クラスターを形成する。この構造体は、星状体(aster)と呼ばれている。前期には核小体は小さくなり、たいていは消失する。また、核膜も消失し各染色分体は動原体のところで紡錘体の微小管と結合する。染色体は移動して、最終的には細胞の赤道面に沿って一列に並ぶ。
●中期
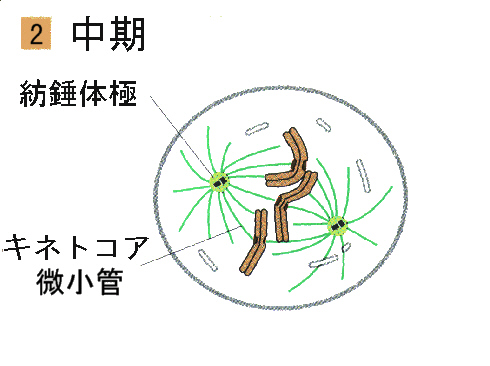
染色体が、細胞の赤道面に沿って並んでいる期間が中期である。この時点で紡錘体は完成している。紡錘体は各極から赤道面に伸びてそこで重なっている微小管と、動原体から極に伸びている微小管からなっている。分裂中期には各姉妹動原体は、紡錘体微小管で両方の極に結び付けられている。
●後期
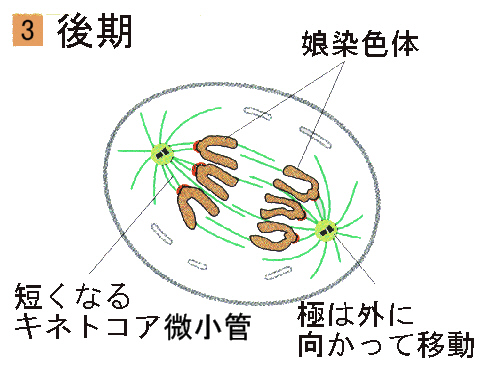
後期は、セントロメア付近で結合していた姉妹染色分体が分離しはじめることにより始まる。それぞれの染色分体はここで独立した染色体となる。分裂した染色体はゆっくりと両極へと移動する。
後期は、全ての染色体が極に到達した時に終了する。重要な過程がこの時期におこっているにもかかわらず、染色体の移動の全体の機構はいまだにほとんど解明されていない。微小管には弾性や収縮性がない。それなのにどのようにして染色体は離れていくのであろうか。染色体は押されるのか引かれるのか、また別の力が作用しているのであろうか。
●終期
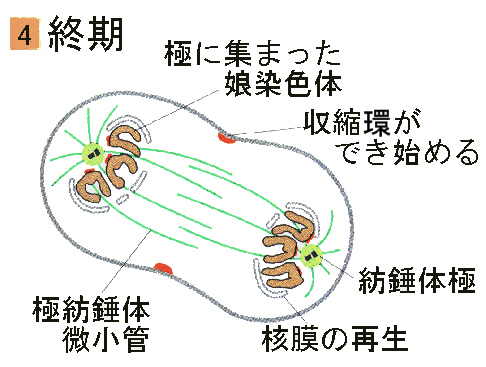
有糸分裂の最終段階である終期は、間期に似た状態に戻る時期である。染色体は、コイルが解けて脱凝縮する。
新しい核膜がそれぞれの染色体のセットの周りに、少なくとも一部は小胞(ゴルジ体由来)や、古い核膜から得られる成分から形成される。紡錘体の微小管は消失し、核小体が見えるようになる。
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
Graphic Gallery
●細胞質分裂(cytokinesis)
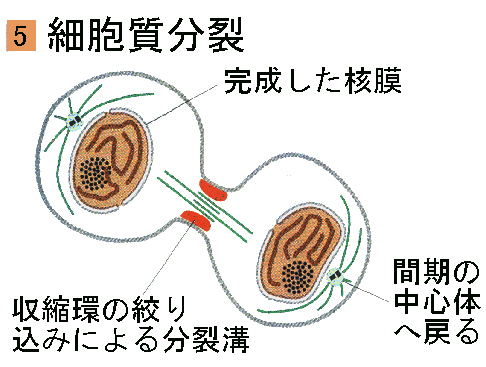
細胞質が分かれて二つの娘細胞が生じる細胞質分裂は、有糸分裂とオーバーラップして、たいていは終期の間に始まる。
動物細胞の細胞質分裂は、細胞が赤道の所でくびれる事で始まる。アクチンとミオシンでできた収縮環が細胞質を絞り込んで、分裂溝がしょうじ、やがて完全にくびれる。
一方、植物細胞では、紡錘体の赤道領域に細胞板が形成され、これが後で細胞壁となることにより細胞質分裂が起こる。細胞板はゴルジ体から生じる小胞から形成される。各娘細胞は細胞膜を形成し、細胞膜の外側の細胞板の側面に、セルロースの細胞壁を形成する。
有糸分裂によって、染色体は(また、存在するならば中心小体)は確実に分配されるが、細胞質のさまざまな小器官はどうであろうか。
細胞小器官は自分自身のDNAを持っていて、すでに存在していたミトコンドリアや色素体、それらの前駆体の分裂によって生じる。ふつうは間期に分裂して、細胞分裂時には分裂しないが、細胞小器官のコピーは細胞内に大量に存在するので、細胞質分裂時に娘細胞に大体、均等に分配される。
細胞分裂の頻度は種によって、また同じ種でも組織によって大幅に異なる。真核細胞の世代期間は普通は長い。中枢神経系のある細胞は、発生の最初の数ヶ月で分裂を止めるが、一方、血球形成細胞、消化管細胞、皮膚細胞は生きているあいだ、ずっと頻繁に分裂する。細胞周期の長さは、外的刺激との相互作用を通じて細胞に内蔵された正確なプログラムを実行するのにかかる時間である。
真核生物の子孫の残し方には、無性生殖(asexual reproduction)と有性生殖(sexual reproduction)の二つの基本型に分ける事ができる。無性生殖では、一つの親が分裂し、あるいは出芽し、またあるいはバラバラになって、二つ以上の個体を生じる。無性生殖のほとんどの場合、すべての細胞は有糸分裂によって生じるので、その遺伝子の特性は親と同一である。このような遺伝的に同一な生物の集団をクローンと呼ぶ。
これに対して、有性生殖では、二つの特殊な性細胞すなわち配偶子(gamate)がつくられ、これが結合して接合子(zygote)となる過程を含んでいる。たいていは、配偶子は二つの別の親から得られるが、一つの親が両方の配偶子をつくることもある。動物では、卵(egg)と精子(sperm)が配偶子で、受精卵(fertilized egg)が接合子である。
もし、配偶子が親細胞と同じ数の染色体をもっていると、接合子は二倍の染色体を持つ事になり、この倍化は世代ごとに起こってしまう。生物はどのようにして、染色体数が増え続ける事なく、接合子を生産しているのであろうか。有性生殖をする生物が、断続する世代において、一定の染色体を保っているのは、減数分裂(meiosis)と呼ばれる特殊な細胞分裂によって染色体の数を半減させているからである。
高等動植物の体細胞に見られる各染色体は、ペアになっていて、この二本を相同染色体という。相同染色体は大きさ、形、セントロメアの位置がよく似ているばかりでなく、特殊な技術で染色してできる特徴的なバンドパターンも同じである。つまり、体細胞は2セットの染色体をもっていて、1セットは卵から、もう一セットは精子から受け取っているのである。相同染色体では、対応する位置には同じ遺伝的特性を支配する遺伝子が載っている。
細胞や核が2セットの染色体を持っていれば、それは二倍体(diploid)と呼ばれる。1セットだけ持っていれば一倍体(haploid)である。精子と卵子は一倍体で、受精するとそれぞれが一倍体の染色体を出し合うので、受精卵(配偶子)では二倍体の染色体数に回復する。細胞や個体が、3セット以上の染色体を持っていれば、それらは倍数体と呼ばれる。倍数体は動物では比較的まれであるが、植物ではごく一般的である。実際、倍数体は植物の進化で重要な因子となってきた。花をつける植物の30~80%は倍数体である。倍数体の植物は、同じグループの二倍体のものよりも、大きく、強く、また商業的に重要である。
2)減数分裂の特徴
動物では、減数分裂は配偶子(精子と卵)の形成されるときにみられる。植物や他の生物では、減数分裂により一倍体の胞子が生じ、さらに胞子が有糸分裂的に分裂して配偶子となる。2つの配偶子が接合すると、核が融合し接合子は再び二倍体の染色体を持つ。
減数分裂の過程は、次に述べる4つの大きな違いはあるものの、有糸分裂とよく似ている。
1)減数分裂は連続した2回の核および細胞質の分裂が行われ、計4つの細胞を生じる。
2)核の分裂は2回連続して起きるが、DNAと他の染色体の成分は、減数分裂の最初の間期に、一度だけ複製される。
3)減数分裂により生じた4つの細胞のそれぞれは、一倍体の染色体を、すなわち相同染色体の一方のみを含むセットを持っている。
4)減数分裂では、両親の遺伝情報が混ぜ合わされるので、それによって生じる一倍体の細胞は、それぞれ現実にはたった一個しかない遺伝子の組み合わせを持つ。
3)減数分裂の過程
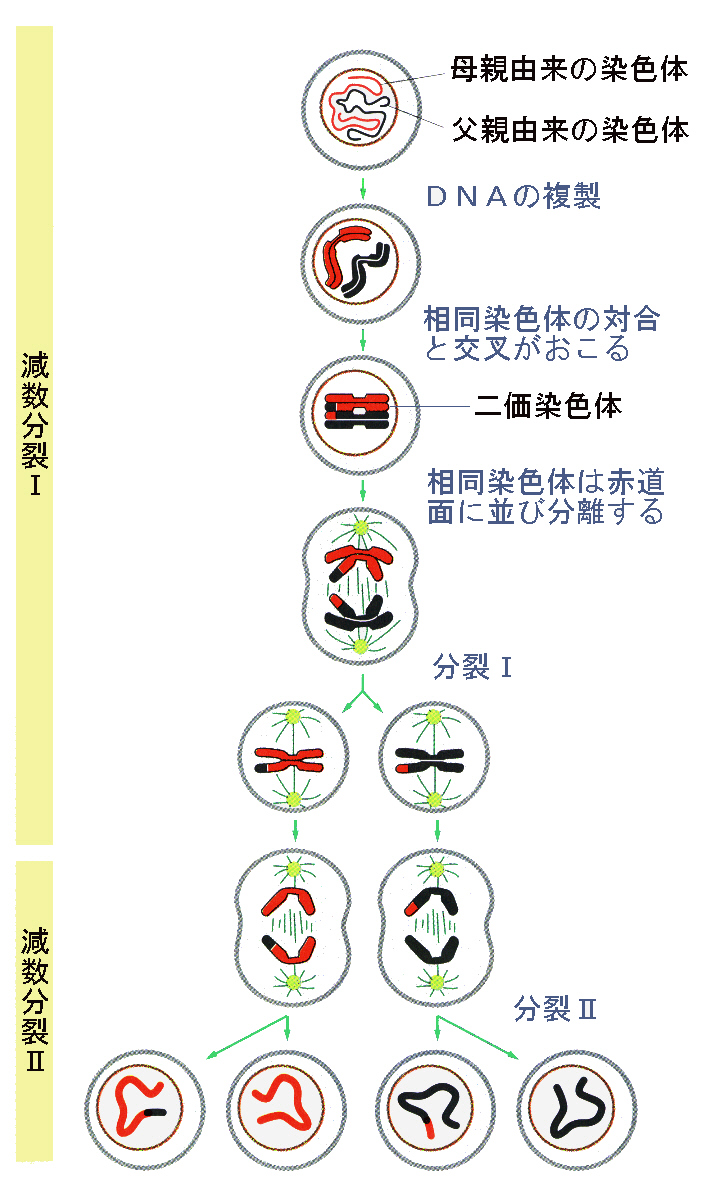
典型的な減数分裂は、二度の核分裂と細胞質分裂からなり、それぞれ減数分裂Ⅰ、減数分裂Ⅱのように書いて区別する。
●前期Ⅰ
有糸分裂と同様に、減数分裂が始まる前の間期のS期に、染色体は複製される。複製された各染色体は、セントロメアのところで結合した二本の染色分体からなる。前期Ⅰの染色分体がまだ細長い間に、相同染色体同士が縦に並び始める。各染色体は二本の染色分体からなるので、対合(synapsis)によって4本の染色分体が一つになる。その結果生じる複合体を二価染色体(vivalent or tetrad)と呼ぶ。二価染色体の数は、一倍体の染色体の数と同じである。相同染色体同士は、対合によってピッタリとくっつく。電子顕微鏡で観察すると、対合した相同染色体の間に、対合複合体として知られる特徴的な構造が形成される事が明らかになっている。このとき交叉(crossing over)によって相同染色体の一部は組み換えられる。この遺伝子組み替え(genetic recombination)によって、有性生殖による子孫の遺伝的な変異が増大することになる。
対合や交叉のようなユニークな過程に加え、有糸分裂の過程に見られたものと似た現象も起きる。
微小管と他の成分からなる紡錘体が形成される。動物細胞では、中心小体の対が両極に移動し、放射状に伸びた星状体が形成される。前期Ⅰの後半には、核膜が消失し、二価染色体の構造が顕微鏡でもはっきり見える。姉妹染色分体はピッタリと結合したままであるが、相同染色体同士はもはやピッタリとはくっついてはいけず、両者のセントロメア(動原体も)はお互いに離れていく。前期Ⅰの後半には、相同染色体は交叉が起こったことを示すキアズマ(chiasma)と定義された特殊な領域の所だけでくっついている。
●中期Ⅰ
前期Ⅰは二価染色体が赤道面に並んだところで終わり、細胞は中期Ⅰに入っている。同一の相同染色体の姉妹動原体は、紡錘糸によって一方の極だけに結び付けられているので、二本の相同染色体の動原体はそれぞれ別の極に結び付けられることになる(有糸分裂ではこれとは対照的に姉妹動原体は両方の極に結び付けられている)。
●後期Ⅰ
後期Ⅰでは、各対の相同染色体は分離し、別々の極へ移動していく。各極は、父親由来と母親由来の相同染色体のどちらか一本だけをランダムに組み合わされたものを受け取る。姉妹染色分体はまだセントロメアで結合している。
●終期Ⅰ
終期Ⅰでは一般に、染色分体はいくらか脱凝縮して核膜が形成され、普通は細胞質分裂が起きる。インターキネシスと呼ばれる間期に似た期間には、もう染色体の複製は行われないのでS期は存在しない。
●前期Ⅱ
多くの場合、染色体は分裂の間も部分的に凝縮したままなので、二度目の減数分裂の前期もまた短い。前期Ⅱでは、多くの点で有糸分裂前期に似ている。相同染色体の対形成はない(実際、各核には相同染色体の対の一方しかない)。交叉もない。
●中期Ⅱ
中期Ⅱでは、染色体は細胞の赤道面に並ぶ。最初の中期と二度目の中期とは簡単に区別する事ができる。中期Ⅰでは、染色分体は4本(二価染色体)に束ねられているが、中期Ⅱでは(有糸分裂と同様に)2本である。
●後期Ⅱ
後期Ⅱでは、動原体で紡錘糸と結合している染色分体が、有糸分裂の後期と同様に引き離され、反対の極へ移動していく。有糸分裂と同じように、染色分体だったものが、この時点で染色体とされる。
●終期Ⅱ
終期Ⅱでは各極には各相同染色体の対の一方が存在している。いずれも複製されていない染色体である。それから核膜が形成され、染色体は次第に伸びて染色質の糸を形成し、細胞質分裂が起きる。
連続2回の分裂は4つの一倍体の核を生じ、それぞれが各染色体の1本を持っている。生じる一倍体の細胞は、それぞれ異なる遺伝子の組み合わせを持っている。この遺伝的多様性は、次の二つの理由による。1)減数分裂では、父方と母方の染色体が中期Ⅰで交叉によって混ぜ合わされる。2)各対の一方が、後期Ⅰで独立してランダムに分配される。
以上の文章は、次の図を参照しながら理解してほしい。
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
Graphic Gallery
最近の研究によって、細胞周期を制御する機構がすべての真核生物に共通であることが分かってきた。
ヒトデの卵成熟を研究していた金谷晴夫は、第一次卵母細胞が停止していた第一減数分裂が再開して卵細胞の成熟開始の指標となる卵核胞崩壊を手がかりに、卵成熟を引き起こす物質を探していた。その結果、1-メチルアデニンという物質が卵成熟を誘起する物質であることを証明した。ちなみにカエルではプロゲステロンが卵成熟誘起物質である。
卵成熟誘起物質は卵内に注入しても卵核胞崩壊を引き起こさない。卵成熟誘起物質が卵外から表面に作用すると、卵細胞質中で最終的には卵成熟促進因子(maturation promoting facotor)が活性化するために卵成熟が開始することが分かってきた。この卵成熟促進因子はその後、すべての真核生物の細胞周期でG2からM期への移行(G2チェックポイント)にはたらいていることが明らかになり、M-phase promoting factortと呼ばれるようになった。
その後、酵母変異株の研究から細胞周期の特定の時期に増殖が停止するCdc2(cell division cycle 2)という遺伝子が見つかり、MPFのもつM期誘起活性の実態は、Cdc2の産物であるCdc2というリン酸化酵素とサイクリンBというタンパク質が結合することによって活性化されたもの(Cdc2キナーゼ)であることが明らかになった。活性化によってM期が開始し、サイクリンが分解されてCdc2と解離することによってM期が終了する。このようにCdc2の活性がG2からM期への移行をチェックしている。
その後、Cdc2と相同性の高い遺伝子がヒトで見つかり、その数が増えていった。これらの遺伝子の産物である、サイクリンと結合してリン酸化酵素の働きを示すタンパク質を、Cdk(cyclin dependent kinases)と総称している。
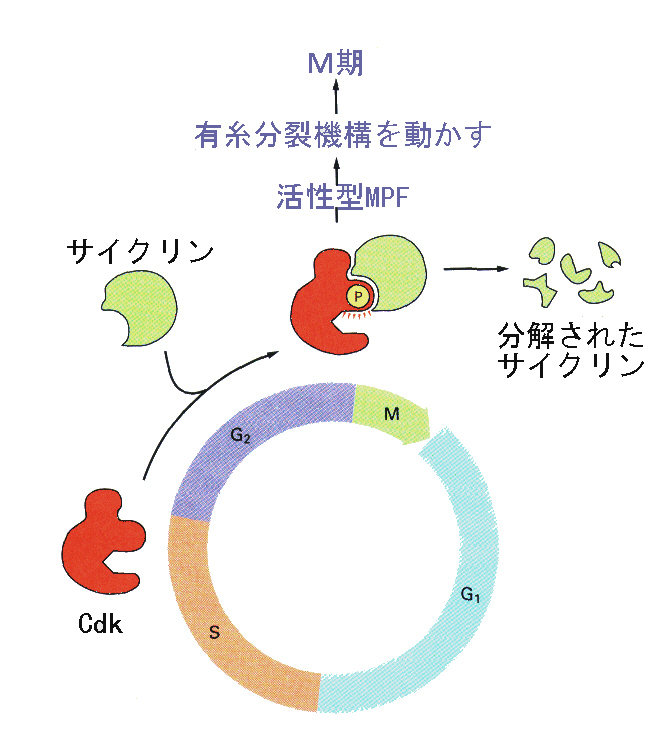
このほか、サイクリン-Cdk複合体、あるいはCdk単体に結合して、タンパク質リン酸化活性を阻害するCdk阻害因子が見つかっている。
細胞周期がスムーズに回転するためには、上に述べたもの以外にいくつかのチェックポイントがあって、常に進行状態を監視している。いずれの場合も、Cdkがエンジン、サイクリンがアクセル、Cdk阻害因子がブレーキの働きをしているとたとえることができる。
さらに学びたい人へ(このページへ戻るときはブラウザーの戻るを選んでください)
http://www.biology.arizona.edu/cell_bio/tutorials/cell_cycle/main.html
![]() この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。