石野研究室
石野研究室
東京医科歯科大学難治疾患研究所・エピジェネティクス分野
〒113-8510 東京都文京区湯島1-5-45tel: 03-5803-4862 fax: 03-5803-4863
(医歯学総合研究棟II期 23階 map)
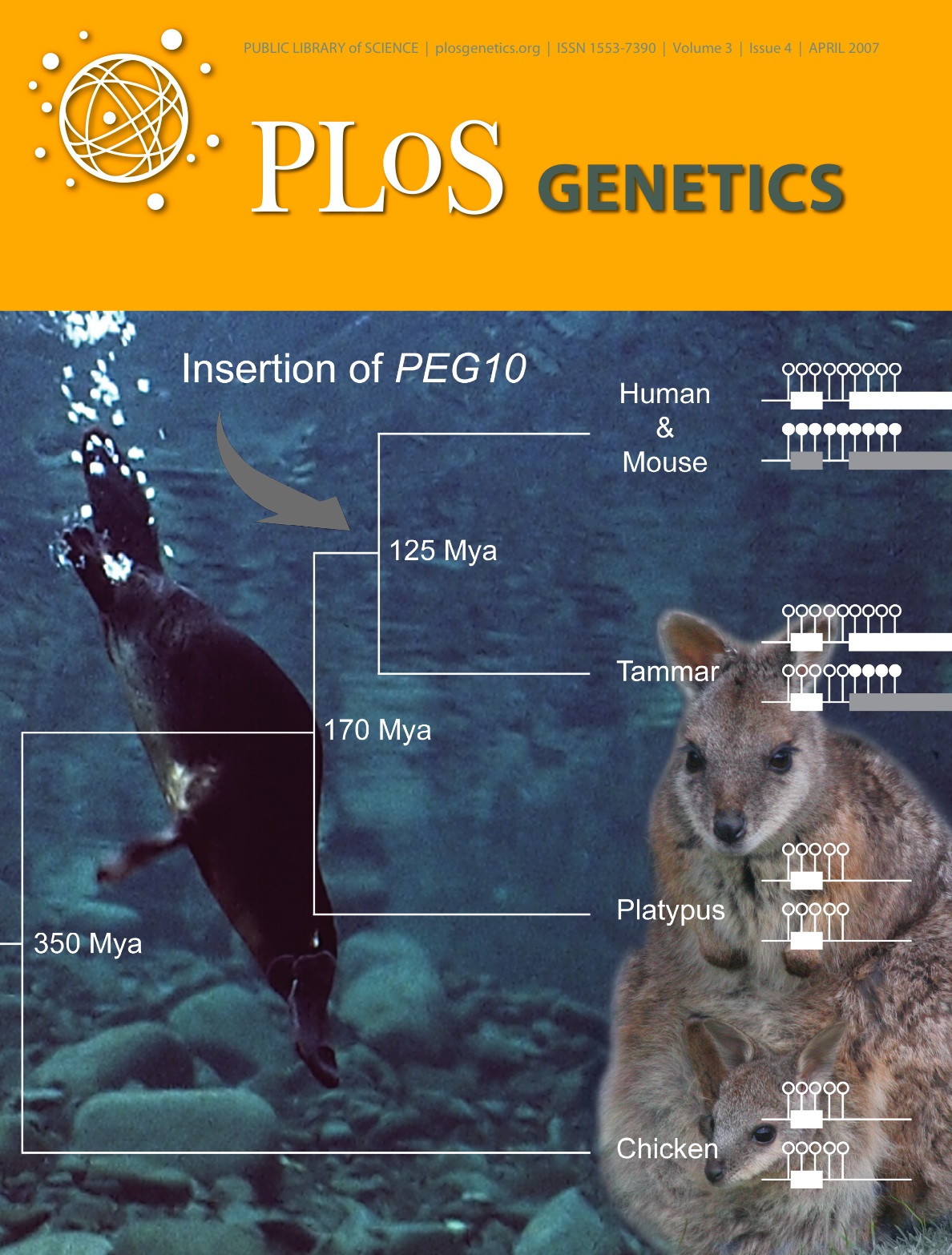
石野研究室マウスにおいて胎盤形成に必須であるPEG10遺伝子は、LTR型レトロトランスポゾンの一種であるSushi-ichiと相同性を有する。したがって、PEG10はレトロトランスポゾンとしてゲノムに挿入された後、進化の過程で胎盤形成に関わる現在の機能を獲得したと考えられる。鳥類や爬虫類にはPEG10は存在しないことがわかっている。哺乳類特異的なこの遺伝子が、哺乳類の進化上どの段階において挿入されたのかは、胎生進化との関係を探る上で重要である。
哺乳類には、マウスやヒトなどが含まれる真獣類、カンガルーやオポッサムなどが含まれる有袋類、カモノハシ、ハリモグラの属する単孔類という3つのサブグループがある。興味深いことに、これらのサブグループはそれぞれ特徴的な生殖様式をもつ。単孔類は卵を産み乳で仔を育てる卵生哺乳類である。有袋類は真獣類と同じく胎生哺乳類であるが、胎盤の機能が低く発生段階の比較的早い時期に仔を分娩し、育児嚢と呼ばれる袋の中で仔を育てる。真獣類は機能の高い発達した胎盤をもち、母体の胎内で長く仔を育てる。
私たちは、これらのグループにおいてPEG10の有無を調べるため、タマーワラビーとカモノハシについてそれぞれの相同ゲノム領域を比較した。その結果、タマーワラビーには高度に保存されたPEG10が存在するが、カモノハシには見つからないことが明らかになった。これは哺乳類が卵生から胎生に進化していく初期の段階においてPEG10がレトロトランスポゾンとしてゲノムに挿入し、哺乳類における胎盤形成の起源に密接に関係した可能性を示唆している。
マウスやヒトにおいては、PEG10周辺は比較的大規模なインプリンティング領域となっている。レトロトランスポゾン由来の遺伝子を含むインプリンティング領域として、ゲノムインプリンティングとレトロトランスポゾンなどの外来配列の不活性化機構との関連性を探る上で非常に興味深い研究対象である。有袋類ではこれまでに、IGF2, IGF2R, PEG1/MESTの3遺伝子についてインプリンティングを受けることが明らかになっている。マウスまたはヒトにおけるこれらの遺伝子は、父親由来、母親由来のアレルでDNAメチル化の異なるゲノム領域、DMR (differentially methylated region)によって発現制御されていることがわかっている。しかしながら、有袋類における相同領域はCpGアイランドになっておらずDMRが見つからないことから、有袋類では真獣類とは異なったインプリンティングの制御機構が進化した可能性が示唆されている。したがって私たちは、PEG10の存在することが明らかになった有袋類における相同領域のインプリンティングの状態を真獣類と比較するため、タマーワラビーのPEG10や周辺遺伝子の発現パターンを解析した。
その結果、マウスにおいてはすべてインプリンティング遺伝子であるPeg10, Sgce, Ppp1r9a, Asb4のうち、有袋類ではPEG10だけがインプリンティングを受けていることが明らかになった。さらに、PEG10の5’領域において、母親由来のアレルのみがメチル化されている有袋類で初めてのDMRを発見した。驚いたことに、真獣類においては隣接するSGCE遺伝子のプロモーターもDMRに含まれるが、有袋類では厳密にPEG10側のみがDNAメチル化の対象となっていた。これらの結果は、レトロトランスポゾン由来であるPEG10がこのインプリンティング領域の起源であり、DMRによるインプリンティング制御機構の共通起源が、真獣類と有袋類が分岐した1億2500万年以上前であることを示すだけでなく、真獣類では一般的であるDMRによるインプリンティング制御機構は、レトロトランスポゾンや外来配列に対するDNAメチル化による不活性化機構を起源としている可能性を提示している。