1.はじめに 2.チャンネルとリンクしていない細胞表面受容体 3.ニューロペプチド 4.動物の行動 5.行動の長期的変容とシナプスの変化 6.脊椎動物の海馬と学習 関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください) 更新日:2001年10月19日
受容体の項で学習したホルモンのはたらきとは異なり、リガンド結合型チャンネルによるシナプスでの信号の流れ(早いシナプス伝達)は、直接的で、単純で、しかも短い時間でシナプス後膜に効果を及ぼす。
これに対して、チャンネルとリンクしていない細胞表面受容体(non-channel-linked receptor)による信号の流れ(遅いシナプス伝達)は、遅く、複雑で、長く続き、場合によっては空間的に広い範囲に効果が及ぶ。
このような遅い効果は神経修飾(neuromodulation)とも呼ばれる。これは同じ細胞上にあるリガンド結合型チャンネルによる早い反応が、チャンネルとリンクしていない細胞表面受容体によって、修飾されているからである。
チャンネルとリンクしていない細胞表面受容体は、ホルモンや局所ホルモン(local chemical mediator)の受容体と、まったく同じような機構で作用をあらわし、実際に受容体は同じである。
チャンネルとリンクしていない細胞表面受容体には、 すでに述べたように、1)リガンドが結合すると細胞内タンパク質のチロシンをリン酸化するようにはたらく、酵素連結型受容体と、2)GTP結合タンパク質を介して膜結合性酵素あるいはイオンチャンネルを活性化する、Gタンパク質連結型受容体に分けられる。チャンネルとリンクしていない細胞表面受容体のほとんどは後者で、Gタンパク質の性質によって次の3つに分けられる。
● Gタンパク質はアデニル酸シクラーゼを活性化、あるいは不活性化し、cAMPのレベルを調節する。cAMPはタンパク質キナーゼA(PKA、Aキナーゼ)を活性化し、多くの標的タンパク質とともに膜のイオンチャンネルタンパク質をリン酸化し、その性質を変える。cAMPは直接イオンチャンネルに結合し、その性質を変えることもある。
● Gタンパク質はイノシトールリン脂質系を活性化し、これがタンパク質キナーゼC(PKC、Cキナーゼ)を活性化し、カルシウムイオンをストアサイトから放出させる。タンパク質キナーゼCはイオンチャンネルをリン酸化し、その性質を変える。さらにカルシウムイオンは、直接イオンチャンネルの性質を変えるか、カルシウムイオン依存性タンパク質キナーゼ(Ca-キナーゼ)がチャンネルをリン酸化してイオンチャンネルの性質を変える。
● Gタンパク質は直接あるいは間接的にイオンチャンネルを開閉する。
いずれの場合も、細胞内メッセンジャー分子が関与しており、情報は細胞内に拡散する。このカスケード反応の段階(ステップ)数が多ければ多いほど、増幅と信号の修飾が容易になる。
50以上の伝達物質が、チャンネルとリンクしていない細胞表面受容体に作用することが分かっている。アセチルコリンのように、チャンネルとリンクしていない細胞表面受容体とリガンド結合型チャンネルの両タイプの受容体と結合して、異なる作用をあらわす例もあるが、多くの場合は片方のタイプの受容体、すなわちチャンネルとリンクしていない細胞表面受容体と結合する。
チャンネルとリンクしていない細胞表面受容体の関与する反応は、数百ミリ秒かかる遅い反応であり、しかも信号分子は拡散して、広い範囲に作用する。
たとえば、平滑筋に神経終末をつくっているノルアドレナリンを放出する神経を考えてみよう。平滑筋は横紋筋とはだいぶ異なるので、まず平滑筋の特徴から述べていく。
1)平滑筋について
平滑筋は血管、消化管、膀胱、子宮など、いわゆる内蔵の器官の管壁を構成し、血管の太さの調節、消化管運動の調節(蠕動運動)、膀胱や子宮などの泌尿生殖器の機能の調節、瞳の大きさの調節など、多くの生体反応に重要な役割をはたしている。
ヒト十二指腸の平滑筋(http://www.med.toho-u.ac.jp/anat1/anatomy/t13.html)より
平滑筋は、骨格筋と違って不随意筋で、自律神経の支配を受けている。
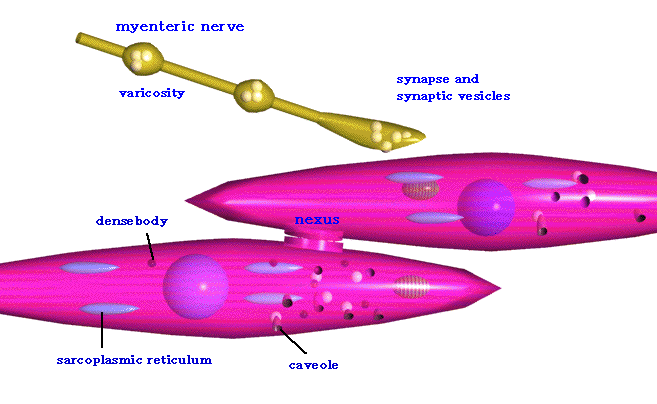
平滑筋には収縮要素であるアクチンとミオシンの規則的な配列がないので、横紋が見られないが、アクチンとミオシンを含んでいないわけではない。アクチンとミオシンは、中間径フィラメントによって細胞膜と結びつけられている。Z膜もT管系もないが、Z膜の代わりにdense bodyによってアクチンフィラメントの端が束ねられている。
また平滑筋細胞同士は、ギャップ結合(nexus)によってお互いに連絡している。
薬理学電子教科書(http://pharma1.med.osaka-u.ac.jp/textbook/SmoothMuscle/SmoothMuscle.html)より
平滑筋の収縮の開始が、カルシウムイオンによっておこる点は横紋筋と同じだが、その他の点はだいぶ異なっている。
● 筋小胞体の発達は悪く、カルシウムイオンは筋小胞体からも放出されるが、大部分はカルシウムチャンネルを通って細胞外から流入する。
● アクチンとミオシンの結合による収縮は、骨格筋ではアクチンに結合したトロポミオシン、トロポニンによっているが、平滑筋にはトロポニンは存在せず、ミオシンの方を修飾する。
カルシウムイオンはカルモジュリンと結合し、カルシウムイオン-カルモジュリン複合体となって、ミオシン軽鎖キナーゼを活性化する。このキナーゼはミオシンをリン酸化して、アクチンと結合できるようにする。こうして収縮運動が始まる。● 横紋筋では、アセチルコリンによって電気的な変化がおこり、その結果、カルシウムイオンが筋小胞体から放出されるが、平滑筋では、交感神経系のアドレナリンと、副交感神経系のアセチルコリン(ムスカリン様)の二重支配を受け、細胞内メッセンジャーがチャンネルタンパク質を修飾して、カルシウムイオン濃度が調節される。
このような特徴があるために、横紋筋の素早い収縮(0.1秒)と異なり、平滑筋の収縮はゆっくりとした(数秒から数十秒)収縮過程をたどり、収縮の加重がおこりやすい。横紋筋の位相性筋に対して、緊張性筋と言う。
さらに学びたい人へ
http://www.ursa.kcom.edu/LectStreams/Other/DesMoines/SmMuscle_DesMoines_files/frame.htm
http://pharma1.med.osaka-u.ac.jp/textbook/SmoothMuscle/SmoothMuscle.html
2)消化管平滑筋の神経支配
それでは、消化管平滑筋を支配する神経を見てみよう。
消化管を支配している交感神経系は、消化管のはたらきを抑制し、副交感神経は増強する(rest and digest)。心臓に対するはたらきとは逆になっている。
この神経は、神経繊維の所々に神経膨隆部(varicosity)と呼ばれる膨らみがあり、シナプス小胞を含んでいる。神経膨隆部は横紋筋の神経筋接合部に見られる終板のような特別な構造をつくらず、シナプス後膜にぴったりと接しているわけでなく、放出された伝達物質は広い範囲に作用する。
交感神経系による平滑筋の収縮・弛緩は、チャンネルにリンクしていない受容体によっておこり、細胞内シグナル伝達系を介してイオンチャンネルの活性を修飾しておこるので、ゆっくりとした反応である。
すでに述べたように、交感神経系のアドレナリンと副交感神経系のアセチルコリンの二重支配を受けているが、それだけではなく、他の分子の支配も受けている。たとえば、子宮平滑筋には、オキシトシン受容体があり、脳下垂体神経葉のオキシトシンによって収縮する。このように、平滑筋はいろいろな生物活性物質の影響を受ける。
中枢でも多くの場合、このような傍分泌(paracrine)による放出が起こっていると考えられている。したがって平滑筋で見られるように、いろいろな生物活性物質の修飾を受ける。
3)モノアミン
アドレナリンやノルアドレナリンはモノアミンと呼ばれ、脊椎動物でも無脊椎動物でも重要な伝達物質であり、神経系に広く分布する。哺乳類ではアドレナリンは橋にある青斑核に多く存在する。青斑核にあるニューロンは、その神経繊維を大脳に送っている。このように、モノアミンは中枢神経系で、神経活動を修飾している。
モノアミンには、アドレナリン、ノルアドレナリン、ドーパミン、オクトパミン、ヒスタミン、セロトニンなどがある。
前回、アドレナリンの受容体に関して説明したが、アドレナリンもアセチルコリンと同じように、多様な作用を示す。これは受容体にサブタイプがあるからで、前回説明したのはβであったが、これとは別にαがあり、それぞれがさらに2つに分けられている。
さらに学びたい人へ(血管の収縮機構発見物語、おもしろい)
http://www.interq.or.jp/kyuushu/dgoto/nitric.html
アドレナリンが副腎髄質から分泌されるホルモンでもあるのは興味深い。
モノアミンの一つであるアドレナリンだけでなく、信号分子として使われているような分子は、ニューロンによっても使われていることが多い。特に血圧の維持、消化酵素の分泌、細胞の増殖などのコントロールに使われている比較的小分子のペプチドホルモンや局所ホルモンではその傾向が強い。
神経系で発見されたこのようなペプチドをニューロペプチド(neuropeptide)と呼んでいる。たくさんのニューロペプチドが見つかっていて、平滑筋のところで述べたようなやり方で、痛みや、快楽、空腹、性などの衝動や気分を調節している。
ニューロペプチドは細胞体で合成され、速い軸索流により4日あるいはそれ以上かかって、軸索末端に運ばれる。多くの場合、大分子の前駆体が最初に合成され、酵素的に切断されてできあがる。
また、一つの前駆体から複数のニューロペプチドが作られることが多い。ニューロペプチドを含む顆粒はシナプス小胞より大きく、電子顕微鏡で観察すると、中身が黒く染まる有芯顆粒(cored granule)である。
ニューロペプチドを放出する多くのシナプスでは、神経伝達物質も放出され、別な作用の仕方でシナプス後膜に働く。たとえばカエルの自律神経の神経節にある神経末端からはアセチルコリンとLHRHが放出される。シナプス後膜には少なくとも3種類の受容体が存在する(Jan and Jan, J. Physiol. 327, 219-246より)。
● 早いニコチニック受容体
● 遅いムスカリニックな受容体
● さらに遅いGタンパク質を介するLHRHの受容体
である。
アセチルコリンはコリンエステラーゼによってすぐに壊されるので拡散しないが、LHRHの場合は効果が現れるのが遅いばかりでなく、拡散によって他の細胞にも効果をあらわす。
次に挙げた図は、カエルの交感神経節におけるアセチルコリンとLHRHのはたらきである。
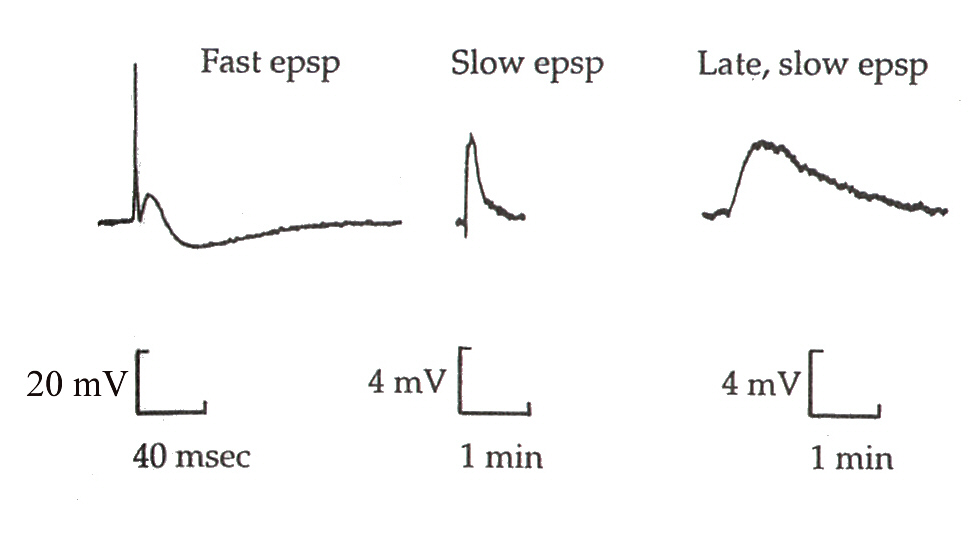
交感神経節の節後ニューロンからは、次の3種のEPSPを記録することができる。それぞれ時間経過と電位の大きさが異なることに注意して欲しい。
Fast EPSPは、アセチルコリンがニコチン受容体と結合したときに生じる電位変化である。一方、Slow EPSPは、アセチルコリンがムスカリン受容体に結合したときに生じる、遅いシナプス伝達による電位変化である。3番目のLate, slow EPSPはLHRHによって生じる電位変化である。
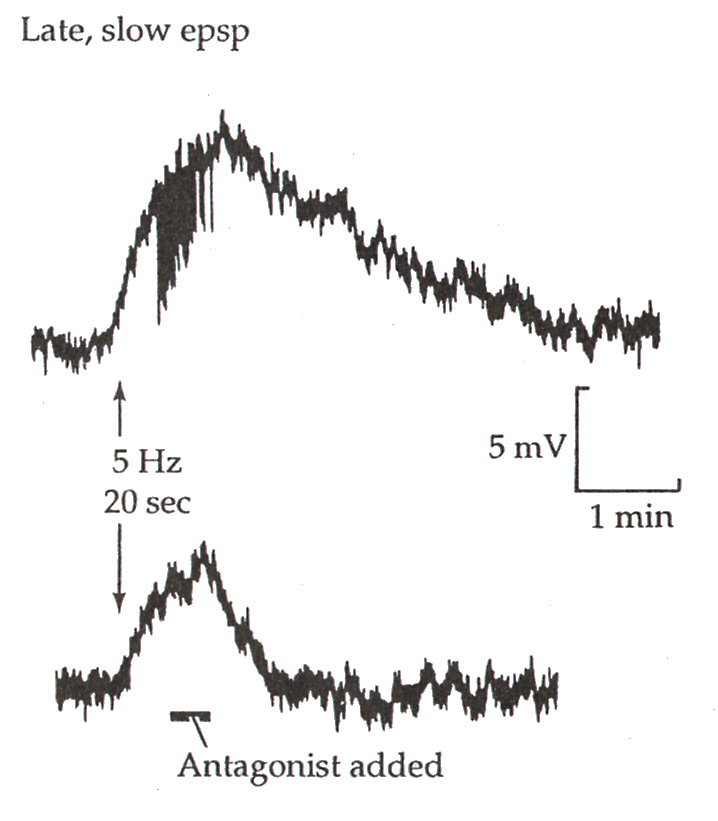
Fast EPSPは、節前繊維を1回、刺激すると、生じさせることができるが、Late, slow EPSPは連続した刺激を繰り返すと、0.1秒後に記録することができ、数分間も続く。
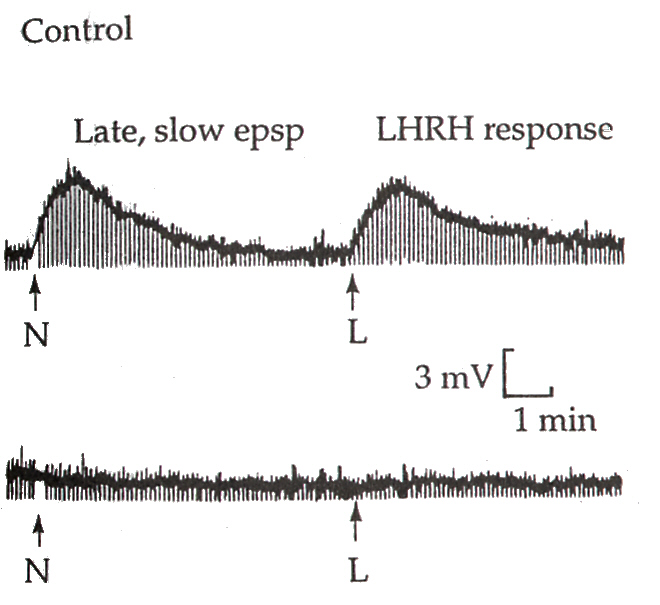
Late, slow EPSPがLHRH受容体を介していることは、次のようにして確かめることができる。下の図のNのところで節前繊維を連続的に刺激した場合の節後ニューロンのEPSPの記録と、Lで節後ニューロンに直接、LHRHを作用させたときの記録を比べてみると、全く同じ電位変化を示していることがわかる。LHRHのアンタゴニストであらかじめ処理をしておくと、NによってもLによっても、電位変化を記録することはできない。
また、下の図のように刺激を加えてEPSPが発生した後にアンタゴニストで処理をすると、EPSPは低下してしまう。
第7,8脊髄神経を刺激すると、Cという節後ニューロンに、Fast EPSPとLate, slow EPSPを生じさせることができ、BニューロンにもLate, slow EPSPが生じる。ところが、第3,4,5脊髄神経を刺激すると、BニューロンにFast EPSPが生じるが、Cニューロンにはなにも生じない。そこで伝達物質と受容体の分布は次のようになっていると考えられる。
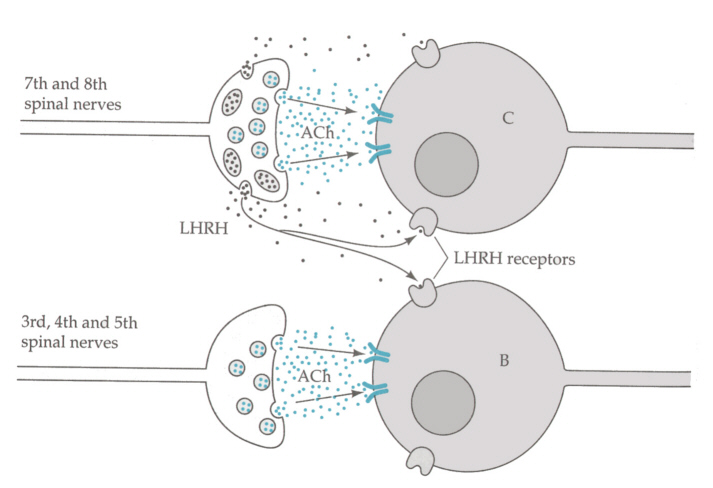
このようにニューロペプチドは近くの他の細胞をも修飾しあうので、はたらきを区別するために分子として大きい必要があったのであろう。脳では、このようなやり方で、さらに広範な神経修飾因子が脳の活動を修飾しているのであろう。
動物は、外界の刺激に対応してさまざまな行動をする。これらの行動は、繊毛や鞭毛、あるいは筋肉によるの運動の総和であり、運動は神経系の支配を受ける。
したがって動物の行動を考えるときには、統御機構の発達を考えに入れないわけにはいかない。統御機構とは中枢神経系のことであり、中枢神経系が発達した動物ほど複雑な行動を行える。
動物の行動は大きく二つに分けることができる。一つは生まれつき備わっている行動、もう一つは学習や知能によって獲得される行動である。
1)生まれながらに備わった行動
ある刺激に対して動物がしめす方向性のある運動を走性という。たとえば光という刺激に向かって進んでいく場合、"正の走光性を示す"という。逆の場合は負の走光性である。光の他にも重力(走地性)、化学物質(走化性)、温度(走温性)、流れ(走流性)などがある。
2)内的行動プログラムと信号刺激
中枢神経系が発達して構成するニューロンの数が増えると、感覚器からの入力および実効器への出力情報を伝えるニューロン以上に、両者をつなぐ介在ニューロンの数が増える。その結果、介在ニューロン間のシナプス結合の数が飛躍的に増大する。
こうしてできた介在ニューロン間のネットワークが、一つの情報処理単位となり、これが組合わさって定型的な行動パターンを生み出す。
中枢神経系には、遺伝子に規定された、このような内的行動プログラムがあって、それが解発されるためにパターン化された行動が起こると考えられている。
よく知られた例として、トゲウオの求愛行動についてみてみよう。
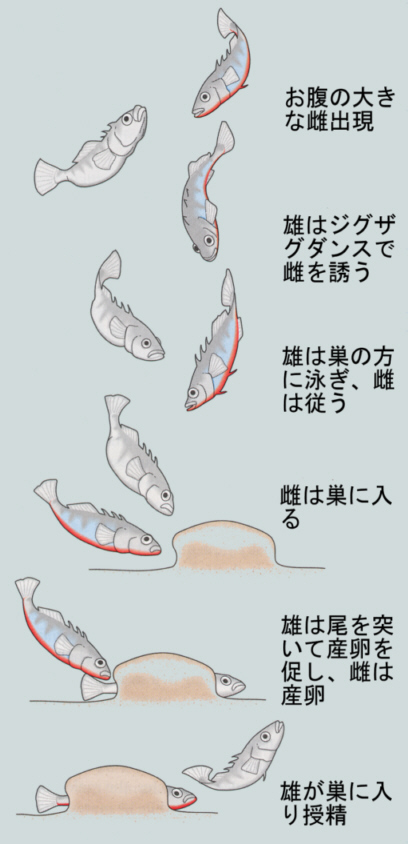
● 繁殖期に入ったトゲウオの雄は、縄張りを構えてその中心に粘液で産卵床を作る。
● こうして雌を待つが、自分と同じ腹側の赤い雄がやってくると、これを追い払う。模型を使った実験によって、追い払うべき雄と認識されるのは下側の赤い細長いものであればよく、トゲウオの形にそっくりでも腹側が赤くなければ効果がない。
この赤い腹のように特定の行動を引き起こす特徴的な刺激を、信号刺激という。
● 一方、卵巣が発達して腹側の大きな雌が近づくと、縄張り雄はジグザグダンスをして雌を誘う行動に出る。
● ジグザグダンスに対して雌が体を反らして反応すると、これが信号刺激となり雄は反応して産卵床へと雌を導く。
● 雌は雄に追随して産卵床に入り産卵し、雄はその後で産卵床に入り精子をふりかけ受精させる。
このような複雑な繁殖行動も、信号刺激を検出する鍵刺激検出機構と、それに対応する特定の行動が繁殖行動の要素の数だけあって、それがリレー式に順序よく解発されて行動が完結するものと考えられる。
下の図は、信号刺激に対して特定な行動が解発される機構を模式的に描いたものである。感覚器からの信号刺激は、鍵刺激検出機構によって受容され、入力中に鍵刺激が含まれていれば、中枢の運動プログラム発生器(CPG)へ送られ、定型的な行動パターンが起こるように運動ニューロンへの出力され、効果器である筋肉が収縮して行動が起こる。
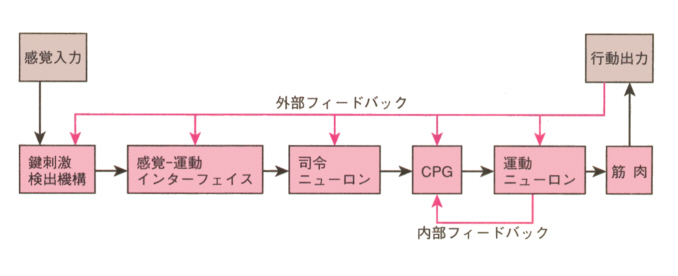
トゲウオの例をとれば、腹側の大きいことが鍵刺激検出機構で信号刺激と受け取られ、これがジグザグダンス発生のCPGへ伝えられ、行動がおこる。
図にあるように、鍵刺激検出機構とCPGの間には、感覚-運動インターフェイスと司令ニューロンがあって、鍵刺激検出機構で受け取られた入力がCPGへ送られる間に、修飾を受けることができる。
トゲウオのCPGは明らかになっていないが、無脊椎動物にも脊椎動物にも、実際に特定な行動パターンを起こすCPGがあることが示されている。
3)生得的行動の修飾
遺伝的に決まった固定的な行動パターンを、生得的行動と呼んでいる。
内的行動プログラムを担う、それぞれの処理単位は、遺伝的に決められたものである。
それぞれの単位間の結線、すなわち軸索によるシナプス結合のできかたは、必ずしもすべて遺伝的に決められたものではない。
神経系が個体発生の過程で、軸索を伸ばしてシナプス結合を作るときには、多めに軸索を伸ばしてネットワークをつくり、結合を形成しなかったニューロンや余分なニューロンは消去している。
このため、ネットワークを形成するときの環境によって、ネットワークのでき方に違いができる。ここに行動が遺伝的な背景だけで決まるのではなく、環境要因によっても影響を受ける理由がある。
このような修飾の例はさえずりの学習に見られる。
ミヤマシトドという北アメリカ産の鳴き鳥には遺伝的に決まったさえずりの鋳型が脳にあって、これを組み合わせてさえずりのレパートリーを形成することが示されている。
雛の時に存在する、この形成過程に全くさえずりを聞かせないで育てると、"下手な"さえずりしかできなくなり、父親のさえずりを聞かせると、それに似た"上手な"さえずりパターンを形成する。
また、この時期に自分のさえずりを聞かないと、やはり"上手く"さえずることはできない。これらの過程は生まれてから50日までの間におこり、さえずりのレパートリーが"結晶化"されるのだと考えられている。
このように、生得的行動は外界および内的な刺激の質と量によって修飾を受ける場合がある。これは一種の学習で、修飾を受けることのない純粋な生得的行動から、次に述べる学習行動までの間は連続していて、両者を画然と分けることは難しい。ここにのべた修飾は、脳幹と古い脳でおこる。
4)学習や知能によって獲得される行動
上で述べた修飾が、もっとダイナミックにかつ可塑的におこるのが学習や知能によって獲得される行動である。哺乳類になって、爬虫類では古皮質と旧皮質の間を占めていたわずかな領域であった新皮質が、飛躍的に発達して左右の大脳皮質となり、ニューロンの数を増やし、その結果、新しいネットワークを形成できることになった。
それまで脳幹で処理されていた感覚情報は、新しい上位のネットワークに伝えられ、ここで処理され、さらに統合されるようになった。このネットワークの広がりは出力面にも反映され、生得的行動は上位の脳から様々な統御を受けることになる。大脳皮質の連合野を中心とした領域に作られるダイナミックなネットワークの形成と消失が、学習や知能による行動を制御している。
ヒトではこの大脳皮質が著しく発達して面積を拡大したため、表面には溝(いわゆるしわ)が多数あり、このため他の哺乳類と比べても著しい学習能力を獲得した。この発達した大脳皮質を使って、ヒトではコミュニケーションの手段として言語が発達した。言語を操るためには、連合野だけでなく口蓋や舌を操る運動野が発達する必要があったが、何より発声器官である声帯の構造と機能の能力向上が必要であった。
5)行動の可塑性
行動の可塑性はニューロン間のネットワークの作りかえによっておこる。軸索が次のニューロンの樹状突起あるいは細胞体とシナプスを形成すると、固定されてしまうように考えられていたこともあったが、現在では、ニューロン間のシナプス結合はもっとダイナミックに変化しているものだということがわかってきた。
また、シナプスに一過性の刺激を与えると、そのシナプスの伝達特性が長時間にわたって変化する。この性質をシナプスの可塑性と呼んでいる。
このシナプス可塑性は記憶の基礎過程と考えられる。このような行動の長期的変容は、どのようにしておこっているのだろうか。
1)アメフラシでの研究
チャンネルとリンクしていない細胞表面受容体による応答はゆっくりしており、しかも長く続く。ここに、この受容体が行動の調節に極めて重要な意味がある。
Eric Kandelによるアメフラシ(下の写真)の研究によって、このことが次第に明らかになってきた。

http://www.enature.com/fotog/fotog_gallery.asp?fotogID=932より
アメフラシは、サイフォンを触ると鰓を引っ込める。繰り返し触っていると、「慣れ(habituation)」が起こって引っ込めなくなる。
頭を叩いたり電気ショックを与えると、「慣れ」はなくなり、再びよく反応するようになる(「感度の強化、sensitization」)。
「感度の強化」は、電気ショックの大きさにより、数分から数時間続き、簡単な短期記憶に相当する。
電気ショックを毎日続けていると、短期記憶は何週間も続くものに変容する。
この行動の変容は、鰓を引っ込める反射を支配している神経回路内のシナプスの変化に帰することができる。
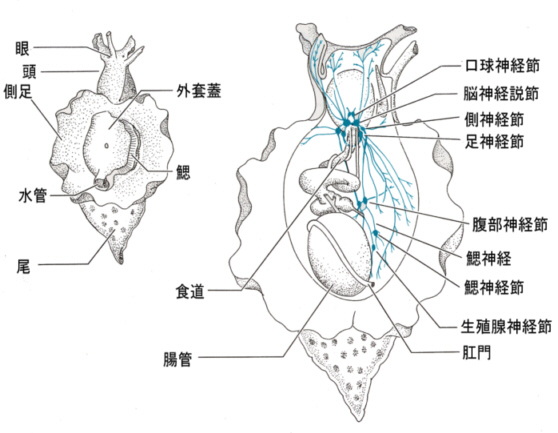
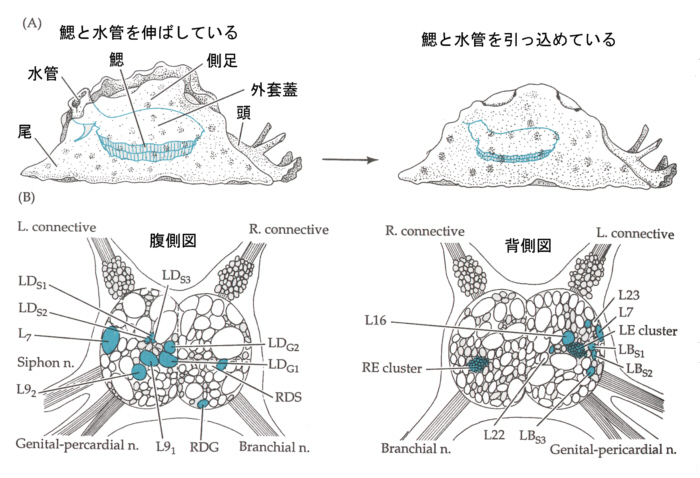
2)アメフラシの神経系
アメフラシのニューロンは大きく(~100μm)、数も少なく(~十万)、行動もステレオタイプなので解析しやすい。
調べて見ると、サイフォンの感覚ニューロンは、腹部神経節にある、鰓を引っ込める運動ニューロンに興奮性シナプスを作っている。「慣れ」のときはシナプス後電位は弱く、「感度の強化」のときは大きくなっている。
この変化は感覚ニューロンから放出される伝達物質の量によっている。したがって、問題は、どうして伝達物質放出量が、シナプスレベルで変化するかということになる。
3)Gタンパク連結型受容体は感度の強化にはたらく
すでに述べたように、シナプスでの伝達物質放出量は、活動電位がやってきたときに、シナプス前膜に流入するCaイオンによって調節されている。したがって、慣れのときはCaイオンの流入が減少し、感度の強化のときはCaイオンの流入が増加すればよい。「感度の強化」について、よく研究されている。
たとえば頭を叩くなどして、「感度の強化」がおこるときには、次の図のようにサイフォンからの感覚ニューロンにシナプスを作る促通性介在ニューロンがセロトニンを放出する。
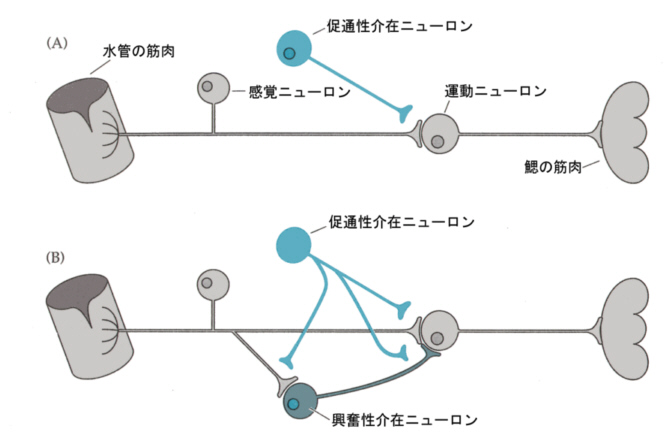
セロトニンは感覚ニューロン上の受容体と結合し、Gタンパク質を介してcAMPの合成を高める。cAMPはAキナーゼを活性化し、Kイオンチャンネルをリン酸化して閉状態にする。すると、静止電位に戻らなくなり、電位依存性Caイオンチャンネルは開状態を続け、そのためにCaイオンの流入が長く続き、伝達物質の放出も多くなる。
これらの実験は、Gタンパク質連結型受容体が、一過性の信号をシナプスの永続的な変化に変え、行動の変容をおこすメカニズムを示唆している。
Kイオンチャンネルの変化は、すぐに脱リン酸化酵素によってもとに戻されるので、短期記憶である。それではどうして、長期記憶が生ずるのだろううか。新しいRNAとタンパク質の合成、あるいは構造的変化を伴う過程がここにはあるのだろうと考えられているが、よく分かっていない。
4)カルシウムイオンとcyclic AMPは無脊椎動物の学習に大切
上に述べたアメフラシでの結果は、ほかの種の記憶や学習にも当てはまるだろうか?
ショウジュバエの突然変異はたくさん知られているが、記憶に関する突然変異に、「知恵遅れ」と「忘れん坊」がある。どちらも学習はするが記憶時間が極端に短い。
変異が起こっているのは、前者がホスホジエステラーゼ(phosphodiesterase)で、後者がCa2+-dependent adenylate cyclaseであった。cAMPが多すぎても少なすぎても記憶形成には不都合らしい。
他に記憶が全く成立しない突然変異体があるが、これはdopa decarboxylaseという酵素が欠損しており、セロトニンやドーパミンを合成できない。
これらの結果は、モノアミンが記憶の最初のステップを担当し、cAMPとCaイオンによるタンパク質のリン酸化が記憶の長期化という次のステップを担っていることを示唆している。
脊椎動物、特に哺乳類では記憶や学習は行動にとって極めて重要である。哺乳類では、大脳辺縁系の海馬が学習に特別な役割を果たしているらしい。海馬(hippocampus)を両側性に破壊すると、古い記憶は残るが記憶の形成がおきなくなる。海馬では連続したインパルスにより長期増強(long-term potentiation)が起こる。
海馬では、シナプス後膜が強く脱分極しているときに、シナプス前膜がインパルスを発射すると長期増強がおこるらしい。この現象にはシナプス後膜の特別な受容体が関係しているらしい。シナプス後膜の脱分極は、通常のリガンド連結型受容体でおこり、グルタミン酸が関与している。
しかしながら、この電位はNMDA受容体と呼ばれるものにより増強される。NMDAとはN-methyl-D-glutamateの略称で、特異的なアナログである。この受容体は、膜が強く脱分極されていて、かつグルタミン酸が結合して、はじめてチャンネルを開く。この受容体が長期増強に関係しており、特異的な阻害剤で阻害すると、シナプス伝達は起こっても増強は起こらない。阻害剤で処理された動物は通常の行動は示すが、海馬が関係すると思われる記憶は形成されない。
この受容体はCaイオンを通すチャンネルで、Caイオンが関係しているらしい(Caイオンの濃度を変えると増強が影響をうける)。
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
http://www.f.u-tokyo.ac.jp/~ikegaya/no0.htm
![]() この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
 ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。
ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。
