 塩基とデオキシリボース
塩基とデオキシリボース1.はじめに 2.DNA分子のつくり 3.糖、リン酸、塩基 4.塩基の配列が遺伝の暗号 5.DNAの複製 6.仁(核小体) 関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください) DNA - The Molecule of Life 更新日:2002/05/02
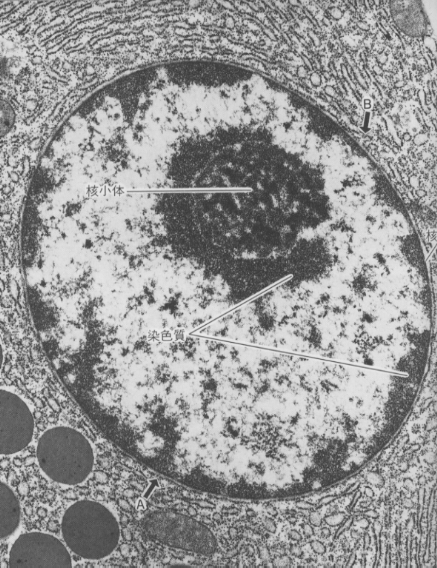
光学顕微鏡で観察すると、細胞分裂をおこなっていない細胞の核の中には、仁(核小体)を除いて特別な構造は見えない。電子顕微鏡を使って高倍率で観察しても、染色質(chromatin)と呼ばれる電子密度の高い部分以外に、特定の構造を見いだすことはできない。ところが、細胞分裂がはじまると、染色質は凝集して染色体(chromosome)が現れる。
染色質が染色体になるのは、体細胞分裂により、間違いなく娘細胞に遺伝情報を分配するためである。したがって細胞分裂のときには、通常の細胞の活動はむしろ一時的に休止している。細胞が生きてゆくために必要な遺伝情報をつねに読み出しているのは、細胞分裂をしていないときである。
それでは、遺伝情報はどのような形で蓄えられているのだろうか、またどのようなメカニズムで読み出されているのだろうか。
遺伝情報はDNAに蓄えられている。
(どのようにして証明されたかは次のサイトを参照してください。キーワード:一遺伝子一酵素説、形質転換(transformation)、ラジオアイソトープで標識したバクテリオファージを使った実験など。
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
http://www.tamagawa.ac.jp/GAKUBU/NOUGAKU/classes/c_biology/biology_o/no3/index.htm
鍵はDNAの構造にある。
ページ先頭へ戻る
DNA(デオキシリボ核酸、deoxyribonucleic acid)は大きな分子(巨大分子、macromoleculeという)である。生体を構成する巨大分子は、多くの場合、単位となるモノマーの重合体である。
●DNA ヌクレオチド
●タンパク質 アミノ酸
●多糖 単糖(グルコース、ガラクトースなど)
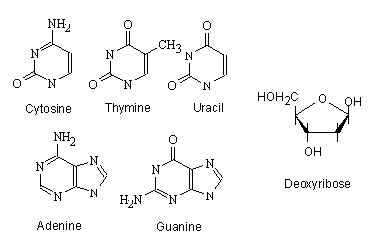
DNAでは、構成単位となるモノマーはヌクレオチドと呼ばれ、リン酸とデオキシリボースと塩基から構成されている。塩基には、シトシン、チミン、アデニン、グアニンの4種がある。
ここにはウラシルも載せてあるが、ウラシルはDNAを構成するヌクレオチドの塩基ではない。ウラシルは、後で出てくるRNAにおいて、チミンのかわりとなる塩基であるが、便宜上ここにあわせて載せておく。
塩基とデオキシリボース
DNAを構成するヌクレオチドは、デオキシリボースの1’(五員環の酸素の右となりの炭素、ここから時計回りに2’、3’、4’と五員環の骨格の炭素に番号を振り、4’から伸びた炭素を5’とする)に塩基が脱水縮合で付き、5’の水酸基とリン酸の間でエステル結合ができた構造をしている。
塩基の種類によって、デオキシシチジン-5’-一リン酸、デオキシチミジン-5’-一リン酸、デオキシアデニン-5’-一リン酸、デオキシグアニン-5’-一リン酸と呼ぶ。
例として、下にデオキシアデノシン-5’一リン酸の分子の構造式を示す。
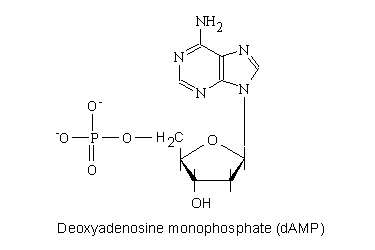 デオキシアデノシン一リン酸
デオキシアデノシン一リン酸
ヌクレオチドの糖部分の3’の位置のアルコール性水酸基は、次のヌクレオチドのリン酸とエステル結合をすることができる。このように次々とヌクレオチドが鎖状に結合して、DNAの片方の鎖ができる。
リン酸を中心にみると、5’と3’の水酸基と2つのエステル結合を作っているので、この結合をフォスフォジエステル結合(phosphodiester bond)と呼ぶ。
例として、下にデオキシアデノシン-5’一リン酸が2つ結合したジヌクレオチドの分子の構造式を示す。
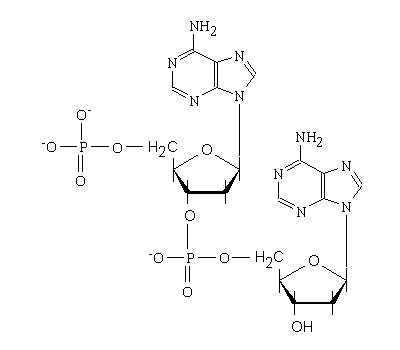 ジヌクレオチド
ジヌクレオチド
たくさんつながったDNAの片方の鎖は、骨格部分が(5’)P-S-P-S-P-S-P-S-P-S(3’)となり(Pはリン酸、Sは糖)、糖の部分から塩基が突き出ている形となる。
塩基は上に述べた4種、シトシン(C)、チミン(T)、アデニン(A)、グアニン(G)である。糖とリン酸の骨格部分はどの部分でも共通なので、違いは塩基の配列の順序になる。そこでうんと省略すると、DNAの片方の鎖はCTAGTCGTAのように書くことができる。このとき約束として5’側を左に書く。
さて、水素原子をもった電子供与基(NやO)が別の電子供与基が向き合ったとき、この水素原子ともう一方の電子供与基の間に水素結合ができる(下図)。
水素結合(hydrogen
bond)は共有結合(covalent
bond)に比べたらもちろん弱い結合力しかないが、たくさん集まればかなりの力になる。その一方で、共有結合に比べたら水素結合を切るエネルギーは小さくてすむ。4種の塩基をみてみると、いずれの塩基もNやOがうまい位置にある。適当な位置で向き合うと、AとTは2本、GとCは3本の水素結合を作ることができることに気がつくだろう。
=O: H-N- → =O---H-N-
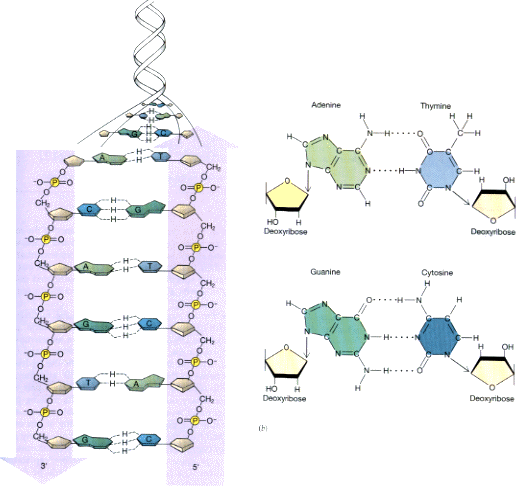
こうして、2本の鎖が1本は5’→3’、もう1本は3’→5’で向き合って、GとC、AとTが水素結合で結合したものがDNA分子である。いずれの塩基も平面的な分子なので、安定した水素結合を作るのに役立っている。したがって、DNA分子は、糖とリン酸の骨格にはさまれて平面的な塩基が積み重なったような構造をしていて、糖とリン酸の骨格はラセン構造をしている。
この構造が非常にユニークな点は、上に述べたように水素結合で2本の鎖が結合している点と、片方の鎖からもう片方の鎖が一意に決まってしまう点である。この性質によって、DNAが遺伝子として選ばれたのである。
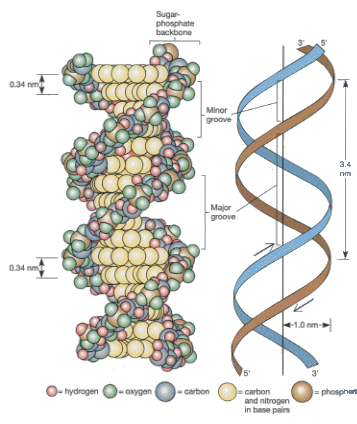
DNAがこのような二重らせん構造をしていることを明らかにしたのは、J. WatsonとF. Crickである。DNAの構造の解明は、分子生物学のブレイクスルーで、これにより多くの未解明な点がおのずと明らかになっていき、さらに多くの研究を誘導した。Nature誌に発表された論文は、わずかなページ数であったが、与えたインパクト極めて大きなものであった。
Nature誌に発表されたオリジナルな論文
http://biocrs.biomed.brown.edu/Books/Chapters/Ch%208/DH-Paper.html
DNAの立体構造(WindowMediaPlayer/4.8M)
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
http://esg-www.mit.edu:8001/esgbio/lm/nucleicacids/dna.html
核には、その生物の全遺伝子情報が存在するが、細胞の特定の機能に必要なモノをつくる設計図はそのごく一部である。遺伝子として機能する塩基の配列部分は必要に応じてほどけ、酵素(RNAポリメラーゼ)の働きによってmRNA(messenger RNA、伝令RNA)の塩基の配列へと転写(transcription)される。
転写の過程(WindowMediaPlayer/10.4M)
たとえば読み取られるDNAの配列がATGGAATTCTCGCTCなら、mRNAの配列はAUGGAAUUCUCGCUCとなる。つくられたmRNAは核膜孔を通って細胞質へ運ばれる。
DNA二本鎖 (5' -> 3') ATGGAATTCTCGCTC (コード鎖、Coding,
センス鎖、有効鎖、sense strand)
(3' <- 5') TACCTTAAGAGCGAG
(鋳型鎖、Template, アンチセンス鎖、antisense strand)
転写された (5' -> 3') AUGGAAUUCUCGCUC
mRNA
遺伝の暗号を解読するために、mRNAを人工的に合成し、これを無細胞タンパク合成系に加え、つくられたペプチドが解析された。根気のいる実験の結果、わかったことは3つの塩基が1つのアミノ酸を指定していることだった。この塩基の3つ組をコドン(codon)と呼ぶ。最初は、暗号表はUCAGを使っていたが、現在では、DNAの塩基に直してTCAGを使うことが多くなった。解読された暗号表は下のとおりである。
暗号表からわかることは、一つのアミノ酸が複数のコドンによって指定されている場合がかなり多いことである。いくつかのアミノ酸では、3番目の塩基が何であってもかまわない。また、化学的に似た性質のアミノ酸は暗号表の近いところにある。
ひと続きのDNAの塩基配列の中から意味のある暗号を読み出すためには、暗号の読み始めと終わりが必要である。暗号表にendと書かれた3つのコドンが終わりを意味する。Metを指定するATGが始まりをも意味する。
| Second Position of Codon | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| T | C | A | G | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| F i r s t P o s i t i o n |
T |
|
|
|
|
|
T h i r d P o s i t i o n |
||||||||||||||||||||||||||||||||||||||||||||||||||||
| C |
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
| A |
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
| G |
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
The Genetic Code
2つの娘細胞には、まったく同じ遺伝情報が伝えられなければならないので、体細胞分裂のときに、染色質が染色体になって、間違いなく娘細胞に分配されるようにしている。そのためには、体細胞分裂の前にDNAは2倍にならなければならないので、体細胞分裂の過程で必ずDNAの複製(DNA replication)がおこる。
すでに述べたように、DNAの構造そのものが、間違いのない複製を保証している。DNAの塩基は、AとT、GとCが相補的に結合するので、片方のDNA鎖に対しては一意にもう片方のDNA鎖が決まってしまうのである。つまり、お互いが鋳型となって、相補的な塩基をつないでいけば、まったく同じ二本のDNA鎖ができてしまうのである。
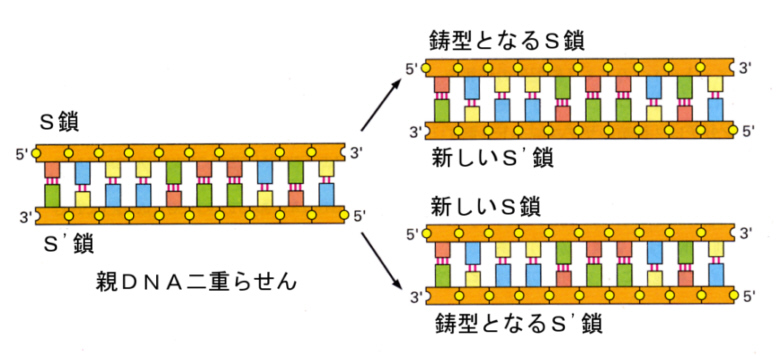
このような複製方法は、「半保存的な」複製と呼ばれている。
それでは、核の中ではどのようにして複製が起こるのであろうか。DNAの複製は、DNAポリメラーゼという酵素を中心とした「複製装置」と呼ぶ多くのタンパク質が関係している。
1)複製開始点
まず初めに、DNAの二本鎖はほどいて、塩基を露出させなければならない。多数の水素結合でしっかり結びついたDNA分子の二本鎖は安定していて、二本を引き離すのは難しいが、一つ一つの水素結合の力は弱い。そのため短いDNAの領域をほどくのには、それほどのエネルギーは必要ない。実際に、複製開始点と呼ばれるDNAの特定な領域があり、ここに楔が打ち込まれるようにしてDNAがほどける。
すると、このほどけた部分に多くのタンパク質が集まって、複製装置を構成する。複製装置は、ほどけた部分から両方向に向かって、次々とラセンをほどきながら移動していく。
ここでもっとも重要なのは、DNAポリメラ-ゼである。この酵素は、ほどけた一方のDNA一本鎖を鋳型として、5’→3’方向に、相補的な塩基を次々に伸ばしていく。付加・伸長の原料となるのは、デオキシリボヌクレオシド三リン酸で、ポリメラーゼはリン酸が2つつながったピロリン酸を切り離し、このとき生じる高エネルギーを利用して反応を進めていく。したがって反応はピロリン酸が切り離される方向、すなわち5’→3’方向にしかおきない。
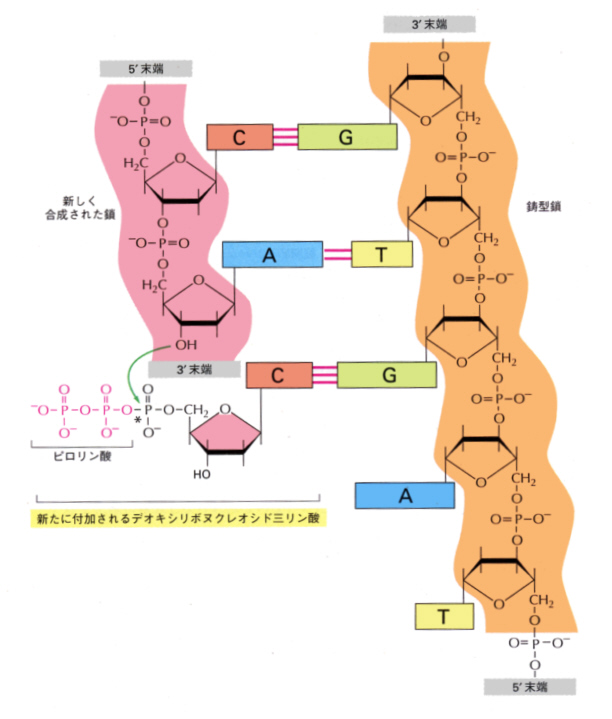
2)リーディング鎖とラギング鎖
DNA鎖は二本鎖で両方の鎖が鋳型となることはすでに述べた。今、上で述べたのはDNAポリメラーゼが複製開始点から次々とDNA鎖をほどいていくときに、5’→3’方向に塩基を伸長させることができる場合について当てはまる記述である。このように素直に塩基の伸長ができる鋳型のほうをリ-ディング鎖と呼ぶ。
それでは反対側の鎖を鋳型にする伸長はどうなっているのだろうか。同じように塩基を伸長しようとすると、塩基の伸びる方向は3’→5’となってしまう。これはDNAポリメラーゼのはたらき方の約束に違反する。
じつは、複製装置にはDNAポリメラーゼがもう一つあって、こちらのポリメラーゼはほどけたDNAのもう一方の鎖を鋳型として、やはり5’→3’方向に短い長さで塩基を伸長すると、ほどけた部分に戻って再び伸長をするという動作を繰り返し、断片を次々と作っていく。このように飛び飛びに伸長せざるを得ないほうをラギング鎖と呼んでいる。
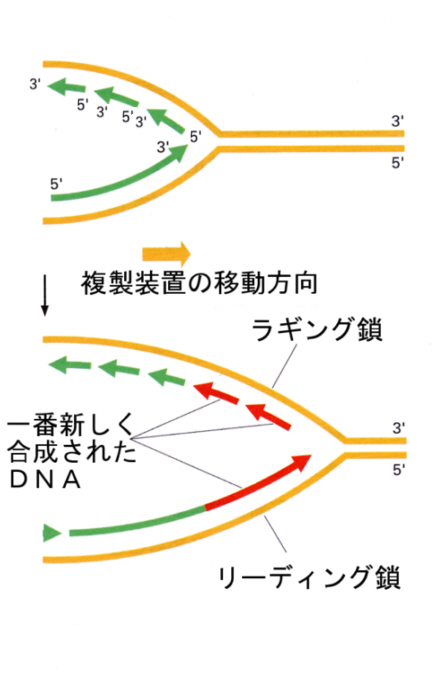
このような飛び飛びの伸長の仕方が、実際におこることを見つけたのは日本の岡崎令二で、今でもこの断片のことを、岡崎フラグメントと呼んでいる。岡崎フラグメントは、あとで繋ぎ合わされて、ラギング鎖でもちゃんとした二本鎖のDNAとなる。
実際には、複製装置は複製開始点から両側に向かって移動していくので、Y字型の複製フォークが向かい合ってつながったような形で、複製が進行していく。また、複製開始点は複数あるので、DNAの複製が始まると、複数の複製開始点から一斉に、上の図で描いたようなことが両方向に進み、短時間でDNAの複製が完了する。
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
http://esg-www.mit.edu:8001/esgbio/dogma/repl.html
http://www.ultranet.com/~jkimball/BiologyPages/D/DNAReplication.html
さらに学びたい人へ(このページへ戻るときはブラウザーの戻るを選んでください)
http://chem-mgriep2.unl.edu/replic/fork.html
http://www.ultranet.com/~jkimball/BiologyPages/D/DNArepair.html
3)校正と修復
実際の複製装置のタンパク質群はもっと複雑だが、ここでは省略する。
複製の過程で誤りの起こる確立は非常に低く抑えられているが、これはDNAポリメラーゼに校正機能が備わっているからで、誤りがあるとそれを正している。
また、DNAに起こった誤りは、そのままにしておくと突然変異の原因となる。そこで細胞には、さまざまの誤りや変異を修復する機構が備わっている(DNA修復、DNA repair)。詳しいことはここでは触れないが、このDNA修復機構が備わっていなければ、生物はこの地球上で生き残ってこられなかっただろうと考えられている。
さらに学びたい人へ(このページへ戻るときはブラウザーの戻るを選んでください)
http://www.nih.gov/sigs/dna-rep/whatis.html
核小体は核の中でも明瞭に区別できる構造である。
ここでは核内で作られたRNAと細胞質で作られ核内に運ばれたタンパク質からリボゾームがつくられる。作られたリボゾームは細胞質に出て、次に述べるタンパク質の合成に使われる。
![]() この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。