1.抗体の多様性はどうして生ずるか 2.細胞性免疫 3.T細胞受容体 4.ヘルパーT細胞 5.ふたたび免疫応答 6.胸腺での訓練 7.自己免疫疾患 8.免疫グロブリンスーパーファミリー 関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください) The Human Immune System 更新日:2001年11月15日
免疫Ⅰの回で、抗体の構造を説明し、抗体がどうやって特異的に抗原を見分けているか、またその見分ける能力の多様性の鍵となる超可変領域について説明した。
免疫系が備えていなければならない性質に、この「特異性」と「多様性」に加えて、「記憶する能力」と「自己と非自己を見分ける能力」がある。あとの2点については後から考えることにして、特異性と多様性がどうして保証されているかについて考えてみよう。
1)抗体の遺伝子
抗体はタンパク質なのだから、遺伝子にその情報が書き込まれているはずである。だとすると、多数の抗原に対応できるクローンを、あらかじめ作っておく遺伝のメカニズムはどうなっているのだろうか。
たとえば100万種類の抗体に対応する100万個の遺伝子を考えなければならないのだろうか。これは長い間免疫学者を悩ませた謎だった。
100万種類の抗体をコードする遺伝子を考えることは、どう考えても難しい。抗体の構造が明らかになり、抗体には定常領域と可変領域があることがわかると、定常領域をコードする遺伝子は一つでよく、可変領域をコードする複数の遺伝子が組み換えを起こせばいいことに気がつく。これだと遺伝子の数が少なくてすむ。
それでも、遺伝子が組み換えをおこして変わるという考えは、遺伝子は突然変異の時以外は変わらない安定したものだという考えに反するものだったのでなかなか受け入れられなかった。
この問題に解決の道をつけたのが利根川進だった(立花 隆/利根川進 『精神と物質』 文芸春秋社、1990)。
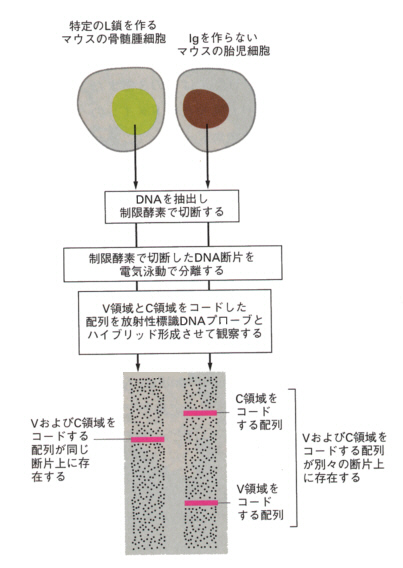
かれは正常マウスの胎児と大人のマウスの骨髄腫細胞から採取したDNAを使い、当時の新しい手法をいろいろ組み合わせて、抗体の遺伝子が胎児のDNAと大人のDNAの間に変化がおこっていることを示したのである。その実験は次の図のようなものであった。
実験には、制限酵素によるDNAの切断、ゲル電気泳動法、ハイブリッド形成(hybridization)などの手法が使われた。ハイブリッド形成とは、一本鎖DNAやRNAの間で、互いに相補的な塩基配列を利用して人工的に二本鎖の雑種核酸分子を形成させることをいう。最初の方法では、ゲル電気泳動後、ゲルの放射能を直接、測定していたが、すぐにサザンブロット法が使われるようになったので、こちらの方法を加えて説明する。
こうして、B細胞の分化の過程でL鎖をコードするDNAの組み換えによる再編成が起こることが明らかになった。多細胞生物では、遺伝子DNAは受精卵から分化成熟する過程で、常に一定不変に保たれているという考えが少なくともB細胞の分化では正しくないことが明らかになった。
マウスのL鎖では、定常領域は単一のCセグメントがコードしている。可変領域をコードしている領域は2つあり、1つは4つのJセグメントが連なった領域、もう1つは多数のV遺伝子セグメントが連なった領域である。
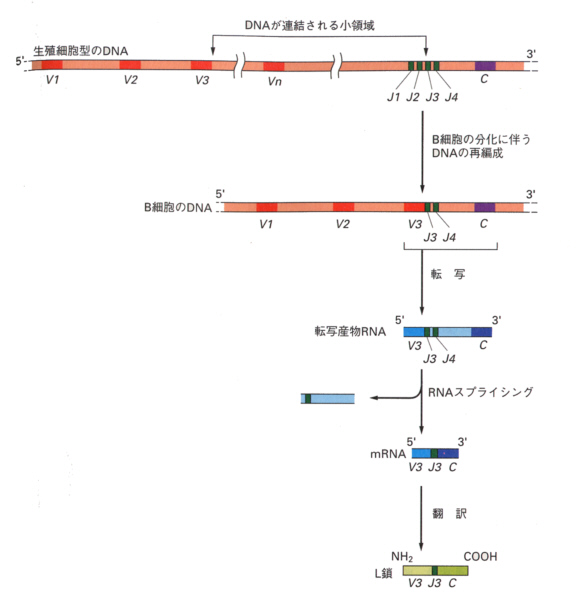
分化の過程でこれらの領域が組み換えを起こし、再編成されてL鎖遺伝子となる。
上の図のように、抗体タンパク質が作られるときは、転写の後、スプライシングによっていらない部分が取り除かれ、V,J,C領域がつながったタンパク質に翻訳される。
H鎖遺伝子はL鎖のV、J、Cセグメントに加えて、少なくとも12のDセグメントが存在する。さらに、Cセグメントは単一ではなく、μ、δ、γ、α、εに対応したセグメントがこの順に連なっている。
大まかな計算をしてみると、L鎖はVセグメントが300あるとすると、Jセグメントが4だから1,200種類のL鎖ができる。H鎖の方はVセグメントを500とすると、500x12x4で24,000の組み合わせができ、L鎖とH鎖が重合すると、1,200x24,000で、2.9x107のオーダーになる。VセグメントとJセグメントの接合部の多様化によってこの数はもっと増え、マウスのB細胞の全数5x108を十分カバーできる数になる。
B細胞は、H鎖のVDJの組み合わせができたあと、それぞれのCセグメントと再編成することによって、生物学的活性が異なるクラスのH鎖を作ることになる(クラススイッチ)。
1)細胞性免疫
ここまでは、体液性免疫とB細胞のかかわりについて話をしてきたので、細胞性免疫とT細胞の関わりについて話を移そう。
やけどをして皮膚が損傷されたので、それを補うために他人の皮膚を移植したとする。移植された皮膚はやがてはげ落ちてしまう。拒絶反応(rejection)である。
拒絶反応がなぜおこるのかを調べることによって、ある個人の細胞には、その個人であることをしめす「表札」のようなものがかかっていることがわかってきた。同じ個体内の細胞にはすべて同じ表札がかかっているが、他人のものとは異なっている。人それぞれ別々の表札を掲げているのである。
この「表札」の本体は膜タンパク質で、組織同士の適合性に関するということから「主要組織適合性複合体(major histocompatibility complex)タンパク質」と名付けられた。これには後で出てくるもう一種類の似たようなタンパク質があるので、区別するためにクラスⅠタンパク質と言う。あまりにも長いので、頭文字を取って「MHCクラスⅠタンパク質」ということにしよう。
他人の皮膚を移植すると、このMHCクラスⅠタンパク質が異なるので、これを手がかりにしてTリンパ球が攻撃を加え、殺してしまうことがわかった。全く同じ作りの集合住宅での玄関で、表札が異なっているので自分の家ではないと認識するようなものである。その後の研究で、MHCクラスⅠタンパク質は、次の図の左側のような構造であることが明らかになった。
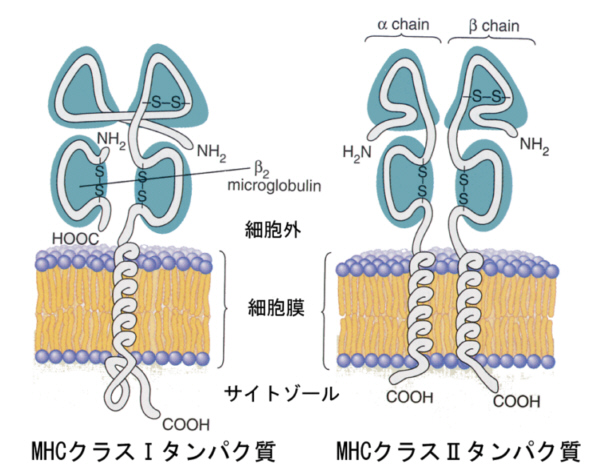
MHCクラスⅠタンパク質の上端の凹みを、上から見たのが次の図である。βシート構造の底にαヘリックス鎖が2本平行に凹みを作っているのがわかるだろう。
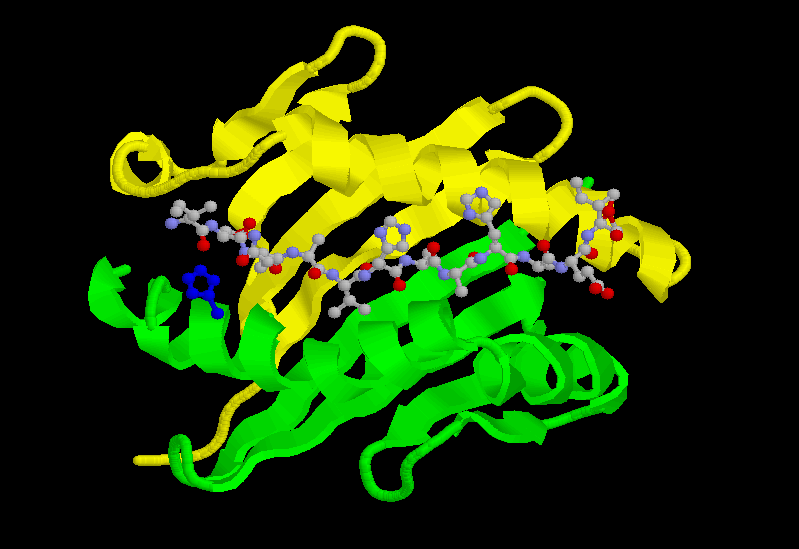
どうしてこのような名前が付いているかというと、MHCクラスⅠタンパク質は、MHC遺伝子群によってコードされるタンパク質だからである。B細胞のところで述べたのと同じように、遺伝子がいくつかの領域に分かれていて、これが再編成されて作られる。このため、上の図の膜タンパク質の上端の凹みの形が、それぞれの個人で異なるのである。
ちなみに、ヒトではMHCタンパク質をHLA(human leucocyte antigen)と呼ぶことも多い。これは、白血球上の抗原として発見されたためである。いろいろな動物で同じような抗原が存在し、移植片拒否に働いていることがのちに分かり、MHCと呼ばれるようになる。
ヒトでは第6染色体の短腕上の動原体近くに接近して存在し、A、C、B(以上クラスⅠ)、DR、DQ、DP(以上はクラスⅡ)の順に並んでいる。複合体といわれるのはそのためである。それぞれが多型を示すので、非血縁の人が同一の組み合わせを持つことはほとんどあり得ない。
このTリンパ球は、異種細胞を傷害して殺すので、細胞損傷性T細胞(cytotoxic T cell、キラーT細胞、CD8+T細胞)と呼ばれる。
2)細胞損傷性T細胞(キラーT細胞)
ここでは簡単のためキラーT細胞と呼ぶことにするが、このリンパ球は異種細胞を拒絶するためだけにはたらくのではなかった。移植というのは自然の状態ではおこらないことである。それではどんなときにキラーT細胞ははたらくのであろうか。
ウイルスなどに感染して、ウイルスが細胞に入り込むと、ウイルスは外皮を脱ぎ捨てる。この外皮のタンパク質の断片が、合成中のMHCクラスⅠタンパク質と結合して細胞表面に現れる。ちょうど「表札」に泥が塗られた状態になる。
細胞損傷性T細胞は、この泥が塗られた「表札」、つまり異種タンパク質断片がはさまった自分のMHCクラスⅠタンパク質を見て、自己ではない(非自己)と認識して攻撃するのである。
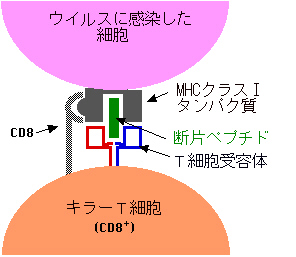
上の図で、CD8と書いてあるのは別の膜タンパク質群(CD=cluster of differentiation)の一種で、キラーT細胞に特異的である。それでキラーT細胞のことをCD8+T細胞とも呼ぶのである。この膜タンパク質は上の図にあるように結合を増強するようにはたらく。
こうしてキラーT細胞が非自己だと認識して結合が強固になると、キラーT細胞は特殊なタンパク質を出してウイルスに感染した細胞を殺すのである。
いままでキラーT細胞は表札を見て、自己の細胞か非自己の細胞かを区別したり、自己の細胞であってもウイルスに感染していて自己とは言えなくなっているのを認識していると書いてきた。
キラーT細胞には「眼」はないので、見ることはできないはずである。どうやって認識するのだろうか。
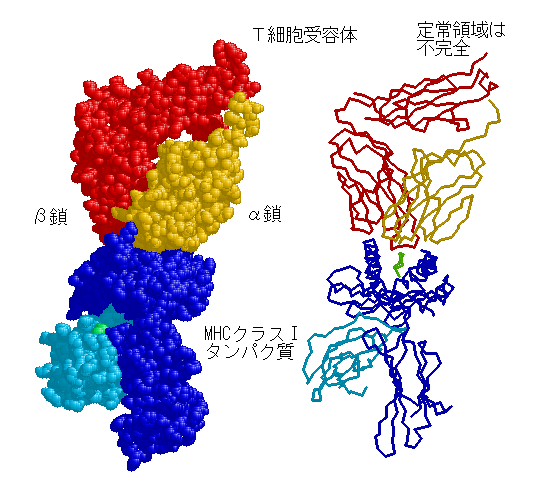
長い間探し求めた結果、1985年になってT細胞の表面には次の図のようなT細胞受容体と言う膜タンパク質があることが明らかになった。
T細胞受容体は、α鎖とβ鎖からなる。どちらも分子量4~5万のタンパク質で、Igと同じようにそれぞれ1個づつの可変領域と定常領域からなっている。可変領域は、やはりIgと同じようにV・D・Jの遺伝子断片の再編成で作られ、α鎖とβ鎖の組み合わせにより複雑な立体構造が作られる。ちなみにα鎖もβ鎖もIgス-パ-ファミリーの一員である。T細胞の受容体は、MHCタンパク質と結合した異物のみを認識する。
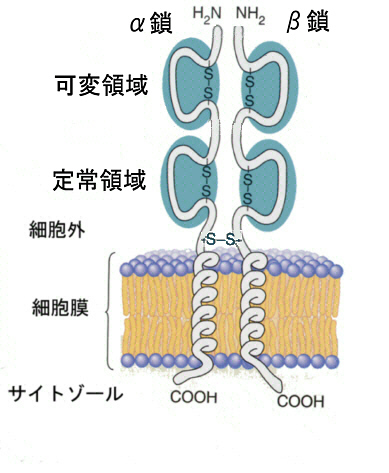
B細胞とまったく同じように、T細胞にもクローンがあって、それぞれのクロ-ンのT細胞受容体の結合部位は異なっている。可変領域の多様化は、抗体の多様性で述べたのと同じような方式で遺伝子の再編成がおこっている。
これまでキラーT細胞は眼を持っていて、これで自己と非自己を見て区別していると言ったが、それぞれのT細胞が見え方の異なる眼をそれぞれ持っていることになる。
ここで、前回出てきたB細胞表面受容体とT細胞受容体の共通点と異なる点をまとめておこう。
B細胞表面受容体とT細胞受容体の共通点:
1)膜に埋め込まれたタンパク質である
2)細胞表面に何千もの同一のコピーが埋まっている
3)抗原に出会う前に、すでにつくられている
4)コードする遺伝子は組み替えによってつくられる
5)特異性の高いユニークな結合部位を持つ
6)この結合部位で抗原のエピトープと結合する
7)結合部位とエピトープの関係は「鍵と鍵穴」の関係である
8)結合は非共有結合である
9)うまく結合が成立し、いくつかの刺激因子があると、
細胞はG0から細胞周期に復帰する
分裂を繰り返しクローンを作る
B細胞表面受容体とT細胞受容体の異なる点:
1)膜タンパク質としての構造が異なる
2)異なる遺伝子がそれぞれのタンパク質をコードしている
3)結合するポリペプチド断片が異なる(表面のエピトープとMHCクラスⅠタンパク質にはさまった断片)
さらに学びたい人へ(TCRとMHCの構造解析のScicenceの総説)
http://sage.bact.wisc.edu/~forest/bact568/humans/tcr.pdf
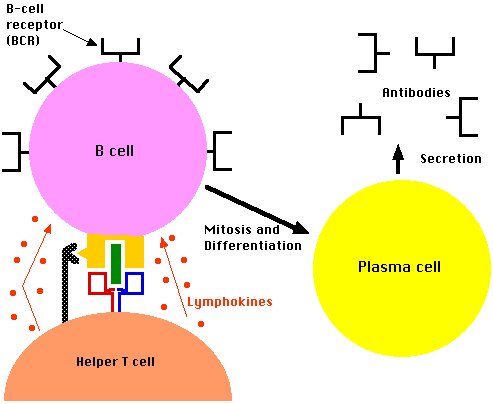
B細胞が形質細胞に変化して抗体を作るようになるのは、B細胞表面受容体と抗原が結合して成熟・分化すると書いた。実は一言ですむような、そんなに簡単な過程ではないのである。
リンパ球の表面にある膜タンパク質がわかるようになり、形態的には区別できなかったB細胞とT細胞がこの表面タンパク質を利用して分けられるようになると、次のような実験が行なわれた。
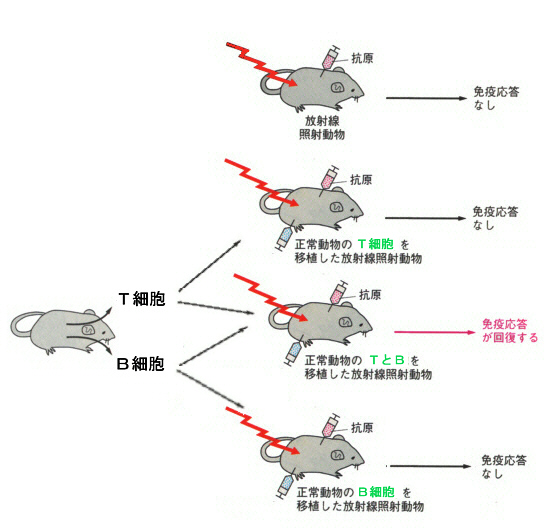
上の実験は、B細胞が抗体を作るようになるためには、T細胞が必要なことを示している。
T細胞には上に述べたキラーT細胞とは異なるヘルパーT細胞(helper T cell、CD4+T細胞)があったのである。キラーT細胞とヘルパーT細胞の違いはCDタンパク質で、ヘルパーの方はCD4をもっている。ヘルパーT細胞もT細胞受容体を備えているのはキラーT細胞と同じである。
B細胞にはB細胞表面受容体とは別に、MHCクラスⅡタンパク質(ずっと上の図参照)を備えている。B細胞表面受容体に結合した抗原は、受容体と一緒に細胞内へ取り込まれる(受容体を介した食細胞運動)。食胞はリソゾームと融合して、取り込まれた中身は分解される。こうして分解された抗原の断片がMHCクラスⅡタンパク質にはさまれて、細胞表面に出てくる。
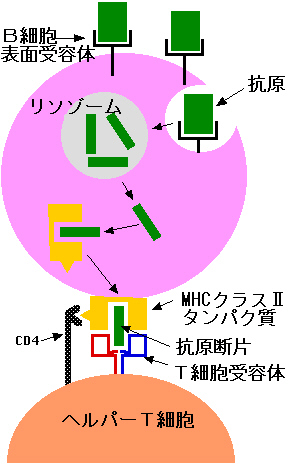
B細胞はこのようにして、非自己成分を細胞表面に提示するのである。そのためこのような細胞を抗原提示細胞と呼んでいる。
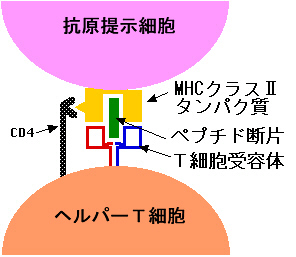
CD4によって両者の結合が強化されると、ヘルパーT細胞は信号分子を出してB細胞に伝える。この信号分子はサイトカインと総称されるタンパク質の一員で、白血球同士の間を取り持つという意味でインターロイキン(IL)と呼ばれる。
インターロイキンはB細胞表面の受容体と結合し、細胞内にセカンドメッセンジャーを作り出し、G0状態にあったB細胞を細胞周期の方へ復帰させる。こうして細胞周期の一連の回転が動き出し、細胞は増殖し分化していく。
1)マクロファージの役割
抗原提示細胞はB細胞だけではない。非特異的な防御機構のところで述べたマクロファージも抗原提示細胞で、細胞表面にMHCクラスⅡタンパク質を備えている。
マクロファージは体内に侵入してきた病原体を貪食するが、その時ただ漫然と食べているのではなく、病原体を分解し、その断片をMHCクラスⅡタンパク質にはさみこんで、細胞表面へ提示する。つまり、「こんな異物が侵入した」と知らせて回るのである。するとヘルパーT細胞がT細胞受容体でこれを読み取り、さらにマクロファージからのILのシグナルを受けて、みずから活性化し増殖する。
こうして活性化したヘルパーT細胞は、多量のILを分泌してB細胞を刺激し、抗体の産生を促進する。
マクロファージやB細胞、ヘルパーT細胞が出会うのは、二次リンパ器官であるリンパ節や脾臓である。ここで接触とILによる複雑な情報交換を行なっている。
2)記憶細胞
B細胞が増殖分化をして形質細胞になるとき、すべての細胞が形質細胞になるのではない。一部は分化していない細胞としてとっておかれる。これを記憶細胞と呼んでいる。
この記憶細胞があるから次に同じ抗原が侵入すると、記憶細胞はすばやく反応して抗体を多量に作ることができる。これを二次反応(secondary response)といっている。
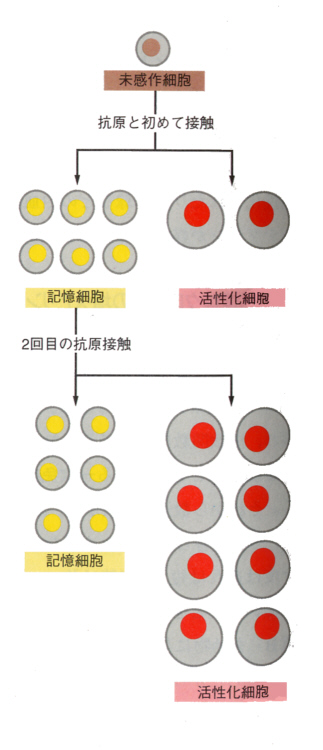
それぞれの抗原に対しては、それぞれ独立した一次反応(primary response)が起こる。
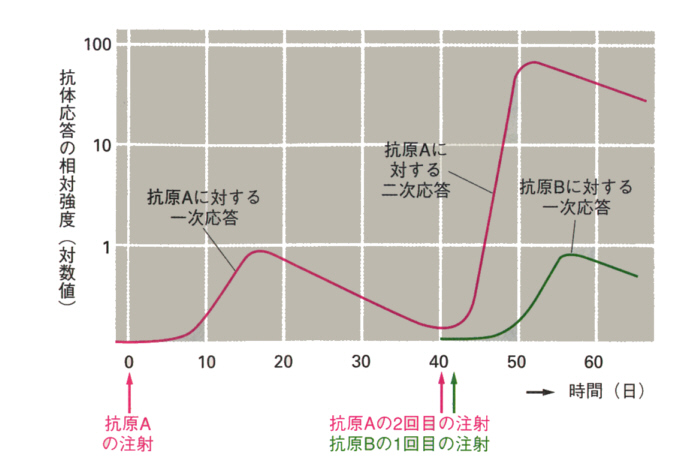
ヘルパーT細胞はシグナル(IL4,5,6)を発して、B細胞を刺激して形質細胞にするか、MHCクラスⅡタンパク質を介して直接接触して刺激する。こうして、B細胞は抗体を生産するようになる。
上の文でILと書いたのはインターロイキン(interleukin)というタンパク質である。マクロファージやT細胞は、IL分子の信号でお互いに交信しあっている。インターロイキンとは、白血球(leucocyte)の間を取り持つ物質という意味でつけられた。ILはもっと広く細胞どうしの信号として働く場合もあるので、サイトカイン(cytokine、細胞を動かす物質)と呼ぶこともある。これらのタンパク質はホルモンと同じように、受容体で受け取られて細胞内にその情報が伝えられる。
まえに、骨髄幹細胞がT細胞になるためには胸腺で訓練を受けるといった。それはどんな訓練だろうか。
生まれたばかりの幹細胞が胸腺に入ると、T細胞抗原受容体の遺伝子が、B細胞の多様化のところで説明したのと同じようにつなぎあわされ、多様な細胞群ができる。これらはメッシュのように発達した胸腺の上皮細胞の上で選別される。ここまでは単に偶然の組み合わせで作られたので必要のない受容体をもった細胞までできてしまうからである。
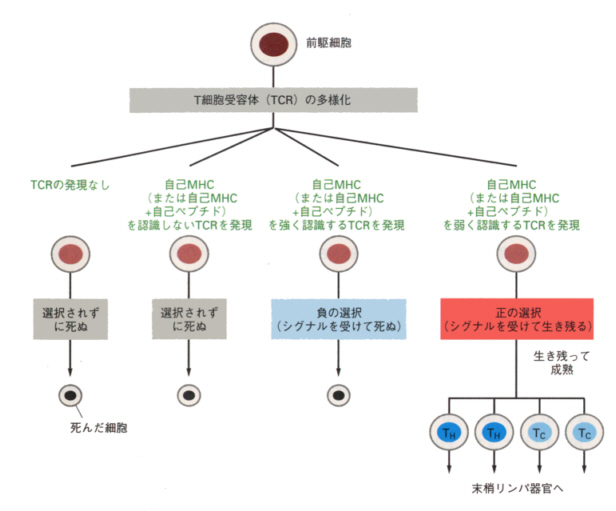
まず最初に、MHCタンパク質を認識できるかどうかが試される。認識できないT細胞は生き延びられずにアポトーシス(プログラム化された細胞死)により死んでしまう。
次に、自己のMHC(あるいはMHCと自己のペプチドの結合したもの)に強く反応するT細胞もアポトーシスをおこして死んでしまう。
こうして、自己が侵害された時のみそれを認識し、自己そのものを破壊することがないことを保証されたT細胞のみが成熟し胸腺から出て、末梢のリンパ組織へ行くことを許される。選択は厳しくて、およそ5%の細胞のみが生き残り、95%の細胞は死を待つ運命にあるという。
胸腺では上に述べた機能を分担するようになり、それに応じて細胞表面分子の発現がおこる。このタンパク質分子がすでに述べたCD(clusters of differentiation)である。CD4分子はヘルパーT細胞、CD8分子はキラーT細胞に特有な分子である。ちなみにエイズウィルスはこのCD4分子に結合し、ヘルパーT細胞に侵入する。
6.で述べたように、自己に対しては免疫機能が働かないように、自己を認識するリンパ球は除去されている。このことを免疫学的自己寛容(immunological self tolerance)と言う。
ところが、これが破れて自己抗原を非自己と認識して、攻撃を加えるようになることがある。このようにして起こる疾患を自己免疫疾患(autoimmune disease)という。
自己免疫疾患には臓器特異性なものと全身性のものがある。その主なものを挙げてみよう。
臓器特異性
血液 貧血、白血球減少症、血小板減少症
中枢神経系 アレルギー性脳炎、脱髄疾患
内分泌腺 慢性甲状腺炎(チログロブリン抗体)、バセドー病(TSH受容体抗体)、
アディソン病(副腎細胞抗体)、糖尿病(インシュリン受容体抗体)、
若年性糖尿病(膵臓ランゲルハンス島β細胞抗体)
消化管 悪性貧血、潰瘍性大腸炎、限局回腸炎
肝臓 慢性肝障害
腎臓 グッドパスター型腎炎(腎糸球体基底膜抗体)、連鎖状球菌感染後腎炎
筋肉 重症筋無力症(アセチルコリン受容体抗体)
眼球 ぶどう膜炎、交感性眼炎(水晶体抗体)
皮膚 疱疹状皮膚炎、紅斑性狼瘡、水泡性類天疱
全身性
全身性エリテマトーデス(SLE)
シェーグレン病
ベーチェット病
リウマチ性関節炎
その他
免疫系にとって完全な自己抗原とは、胎児のうちから一生を通して自分の身体の中に十分に存在し、かつ免疫担当細胞と常に接触することのできる抗原である。
そのために甲状腺のチログロブリン(甲状腺濾胞の中に閉じ込められていて接触できない)や精巣が成熟する思春期になって初めて現われ、しかもリンパ系から隔離されている精子などは、自己として認識されない。
そこでなんらかの理由でこれがリンパ系と接触すると、異物として認識されてしまう。臓器特異性の自己免疫疾患は、かなりのものがこのような発症メカニズムによって理解できる。しかしながら全身性自己免疫疾患については、まだまだ不明な点が多い。
抗体、T細胞受容体、MHCタンパク質、CD4、CD8などはいずれも似た構造をしている。どの分子もアミノ酸70-110個からなるドメインを1個または複数個持っている。ドメインは2つの逆平行のβシート構造がサンドイッチのように向き合い、SS結合で安定化されている。多くの分子は二量体(dimer)かオリゴマーで、一つの鎖のドメインは他の鎖のドメインとインターアクトしあっている。
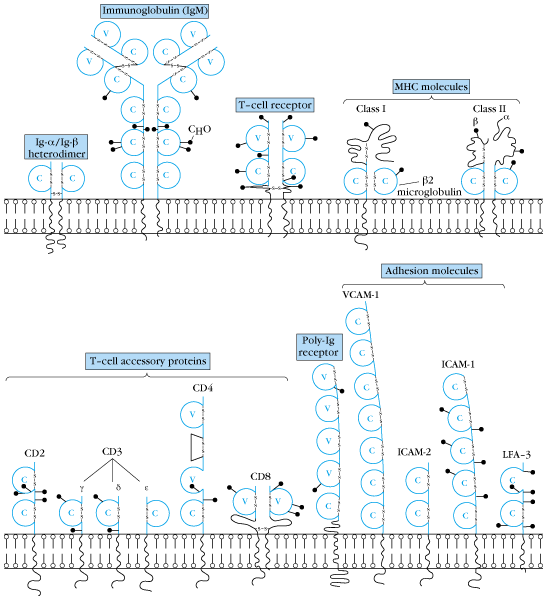
おそらくIg様ドメインの一つから、長い進化の間に遺伝子重複により多種類のファミリーメンバーが生じたのであろう。
![]() この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
 ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。
ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。