
1.発生のしくみ 2.体軸の決定 3.ホメオボックス 4.脊椎動物の場合 5.おわりにむかって 補足 鳥類と哺乳類の初期発生 関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください) http://www.welc.cam.ac.uk/~smithlab/part_1a_lecture_list.htm http://www.med.unc.edu/embryo_images/unit-welcome/welcome_htms/contents.htm http://www.pluto.dti.ne.jp/~sohtsuki/ 更新日:2001年11月22日
1)ごく短い歴史
均一に見える卵が大人の複雑な体制に発生するのを見て、昔のヒトはきっと卵か精子に小さな大人の体制がしまい込まれているのだろうと考えた。発生(development)というのは、それが表に現れてくることに違いないと思ったのである。
顕微鏡を使って精液を観察したAnton van Leeuwenhoekは、精子は種で、卵の畑に植え付けられると考え、精子の中に大人の入れ子を見いだそうとした。同時代のNicolas Hartsoekerは見えたらいいなと思って、実際に子供が精子の中に座っている図まで描いた。しかし、卵の方が重要だと考える人が多かった。
受精という現象が、正しく理解されるようになったのは、19世紀の後半になってからである。その後、無脊椎動物を使って、発生中の胚に対してさまざまな実験的な操作を加えることによって、受精卵が発生の過程でだんだんと大人の体制になっていくことが確かめられていった。その過程で、動物の種類によって、早くから発生の運命が決まっている卵(モザイク卵)と、周囲の影響によってだんだんと決まっていく卵(調節卵)があることがわかった。
やがて、遺伝子DNAが明らかになり、DNAの転写とタンパク質へ翻訳の過程が明らかになり、すべての細胞には、その生物のすべての情報がDNA分子に暗号の形でおさめられていることが明確になった。身体の各部の細胞の形や機能が異なるのは、全情報のうちの必要な部分からだけ転写・翻訳が行われている(発現、expression)からである。
したがって、1個の受精卵から複雑な多細胞の個体になるのは、この発現が時間的、空間的に異なっている結果である。前回の講義で述べたように、発生の最初のスイッチが入ったあと、最初は卵細胞内に蓄えられたmRNAの翻訳がおこり、不活性だったタンパク質が活性化され、発生の過程が進んでいく。その後は、ある遺伝子が発現し、その遺伝情報によって作られたタンパク質が、次の遺伝子の発現を引き起こしていく、といった連鎖反応がおこるのだろうと考えられるようになった。
2)誘導・反応能・分化、拘束(指定・決定)
脊椎動物の眼のうち、水晶体(レンズ)、角膜は外胚葉のうち表皮から、、網膜は神経管から生じる。
神経管の前端に膨らみができ、脳胞ができあがり、3つにくびれるころ、前脳胞の両側から袋状の眼胞がせり出し、やがて前端がややくぼんだ杯状の構造、眼杯となる。眼杯は、厚くなった表皮、水晶体板をくびり取り、表皮は水晶体になる。眼杯の凹みは大きくなり、外側が色素上皮層となり、内側が網膜となる。
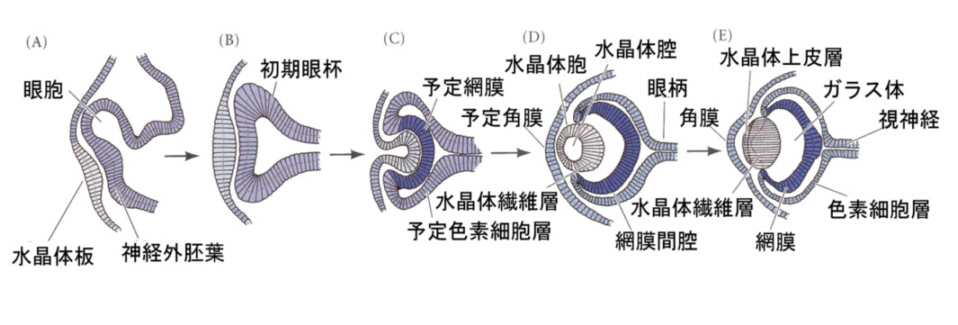
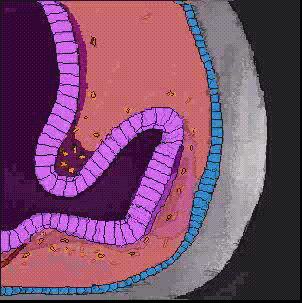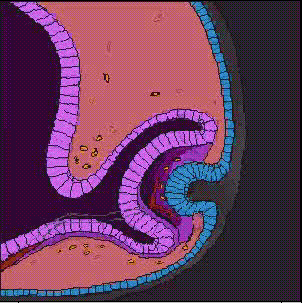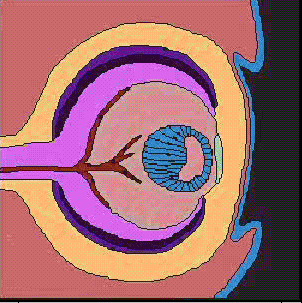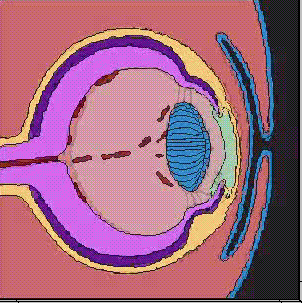
上に述べた眼の発生のとき、表皮外胚葉が水晶体板になり、やがて水晶体胞になるが、これは眼杯が表皮にはたらきかけているためで、このようなはたらきかけのことを誘導(induction)と呼んでいる。この時期に、眼杯を取り除いてしまうと、表皮から水晶体ができてこないことから、眼杯からの誘導があることがわかる。
一方、表皮の方もこの誘導を受け入れる能力があるから、誘導に反応して水晶体に変わる。眼杯を頭部ではない別の部域の表皮の下に移植しても、水晶体は誘導されない。このような受け手の能力のことを、反応能(competence)と呼んでいる。
こうして反応能がある表皮は、眼杯の誘導によって水晶体に分化(dofferentiation)する。分化というのは、表現型として目に見える形で変化したことを言う。
目に見える形で分化が起こる前に、目には見えないが、分化への方向付けが起こっていると考えられる。このような方向付けがおこったことを、拘束(コミットメント、commitment)と呼んでいる。拘束はさらに2つの段階に分けることができる。
水晶体の分化を例に取れば、第一の段階は、水晶体に分化の拘束を受けた表皮を、中立的な環境におけば水晶体に分化するが、他の誘導のもとではその影響を受けてしまうような段階で、このような状態を指定(specification)を受けたと言う。
第二の段階は、水晶体に分化の拘束を受けた表皮を、どんな条件、たとえば他の胚領域において他の組織への誘導の影響を受けても、水晶体に分化するような段階で、これを決定(determination)された言う。
3)種によって異なる卵での指定の時期
モザイク卵と調節卵という語を上に書いたが、動物の種によってその卵の各部分が指定を受ける時期は異なっている。次の3つの型に分類することができる。
●はじめから指定されている(autonomous specification)
多くの無脊椎動物
卵細胞の細胞質に存在する特定の分子の不均一性により指定がおこり、隣の細胞の影響を受けない
モザイク卵という
●条件によってしだいに指定される(conditional specification)
脊椎動物と一部の無脊椎動物
細胞間の相互作用により決まり、相対的な位置が重要
調節卵という
●シンシチウムによる指定(syncytial specification)
昆虫
初めは核だけが分裂して移動するが、移動した先の細胞質によって指定がおこる
しかしながら、この差は本質的なものではない。誘導や指定、決定をおこなうのは何らかの因子があると考え、これらの因子のことを、誘導物質(inducer)とかモルフォゲン(morphogen)と言い習わしてきた。これらの因子は、当然何らかの物質的な基盤があると考えることができる。したがって、誘導や決定の時期の違いは、これらの物質が、最初から用意されているか、後から発現して作用するかの違いだからである。
それでは、誘導、指定、決定、分化はどのようなメカニズムによっておこるのだろうか。
発生を考える上で、軸という概念は極めて重要である。我々の体にはいくつかの軸がある。
前後軸(antrior-posterior axis)=頭尾軸(cepahlic-caudal
axis)
背腹軸(dorsal-ventral axis)
左右軸(left-right axis)
先端基部軸(proximal-distal axis)
何にも無いように見える卵に、どうして体の前後軸や背腹軸が決まるのだろうか。
1)ショウジョウバエの体軸の決定
ショウジョウバエ(fruit fly, Drosophila melanogaster)は1世代の長さが短い(9日、受精してから幼虫として孵化するまでは1日)ので、遺伝の実験には好都合である。染色体の数が少ない(2n=8)のも都合が良い。すでに染色体地図ができているし、多数の突然変異体が維持されている。そこで発生の実験にも盛んに使われるようになった。

関連するサイトとリンク、ショウジョウバエの遺伝情報(このページへ戻るときはブラウザーの戻るを選んでください)
http://flybase.bio.indiana.edu:82/
http://www.pluto.dti.ne.jp/~sohtsuki/
http://dept.biol.metro-u.ac.jp/fly/www/index.html
http://www.dgrc.kit.ac.jp/chromosome/index.html
すでに述べたように、ショウジョウバエの卵割は表割と呼ばれる形式で、最初は核だけが分裂し、それが表面に並び、やがて細胞膜が区画をつくって胞胚ができ上がる。このとき、すでに述べたように極細胞が生じたほうが後部になるので、体の前後軸ができていることになる。
やがて細胞の運動性が増して嚢胚形成が始まり、予定内胚葉、中胚葉、外胚葉が分離する。
まず腹側の正中線をなす細胞群が陥入し、内側に入り込んで管状の中胚葉を作る(下図ステージ7、195-200分後)。この溝を腹溝と呼ぶ。
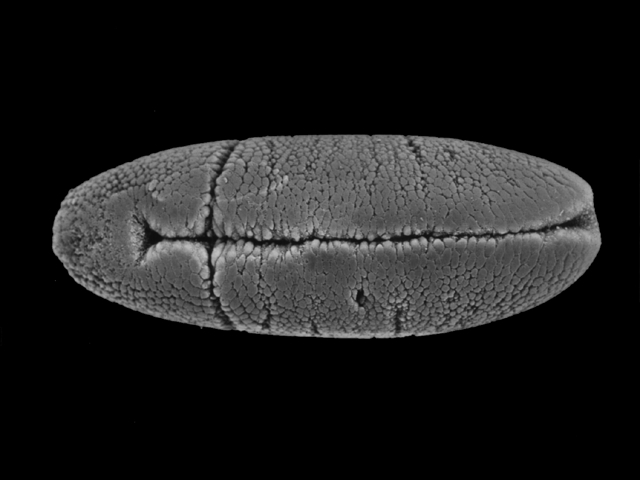
内胚葉は腹溝の前後の端の細胞群が袋状に陥入して形成される(下図上ステージ6、185-195分後)。極細胞はこのとき体内にとり込まれる(下図下ステージ7、195-200分後)。
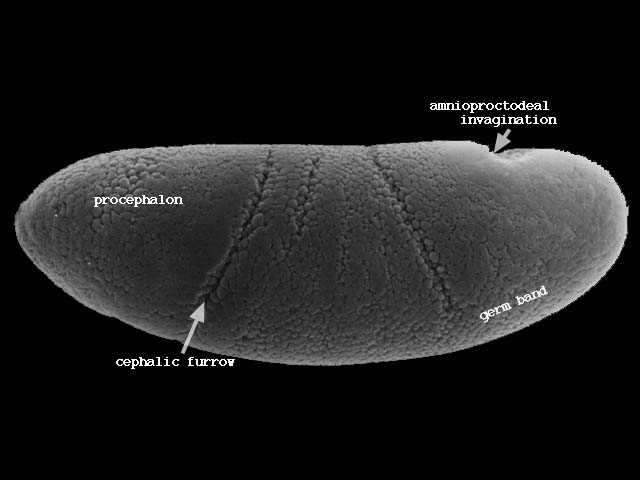
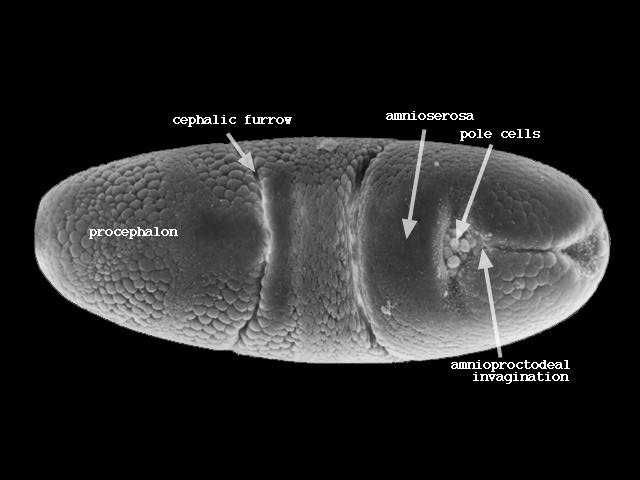
表面に残った細胞群は腹側の中心線に向かって移動し胚帯(germ band)を形成する(下図ステージ8、200-230分後)。
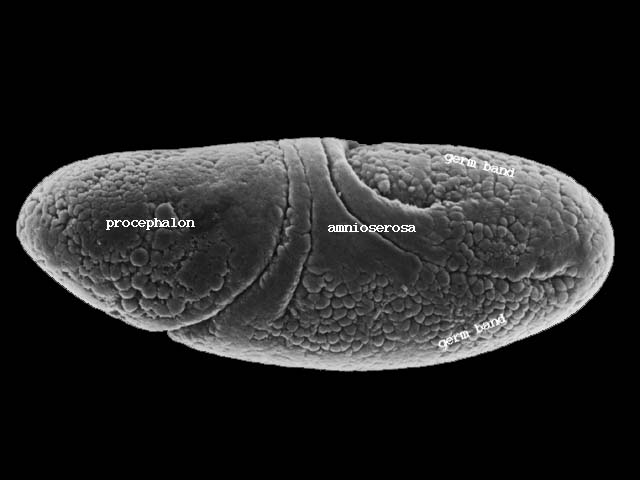
胚帯の細胞はついで後方へ伸張し、さらに後方から回り込んで背側を前方に伸びる(下図ステージ11、320-440分後)。
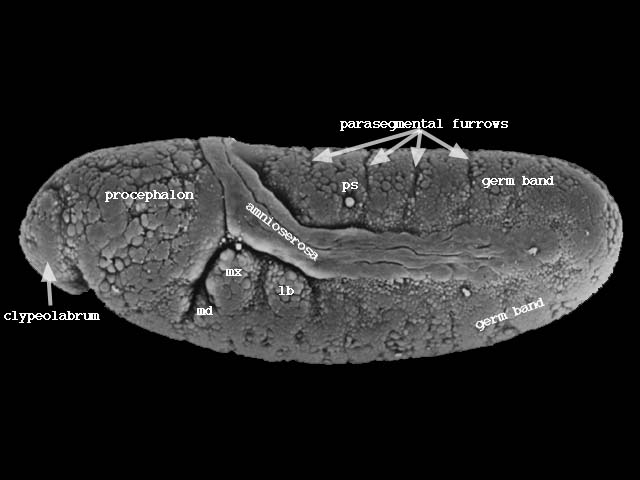
やがて胚帯に体節構造が現れ始める(下図ステージ12、440-580分後)。
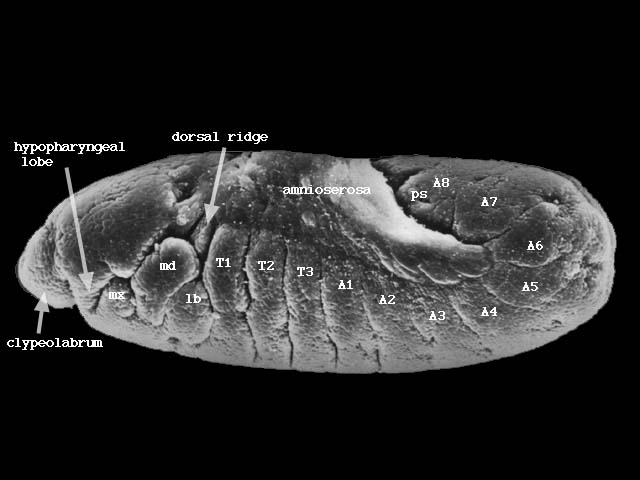
次に胚体は短縮し始め、背部にあった予定後方体節は体の後方に位置するようになる(下図ステージ13、560-620分後)。
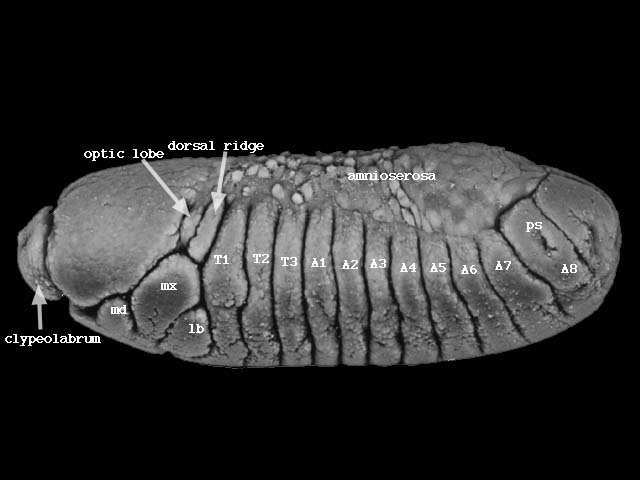
こうして頭部、胸部3節、腹部8節が明瞭になる。それぞれの体節は、体節ごとに発生の運命が異なる。たとえば、胸部第1節は腹側に肢のみ、第2節は背側に羽根と腹側に肢、第3節は背側に平均棍と腹側に肢が生じる。
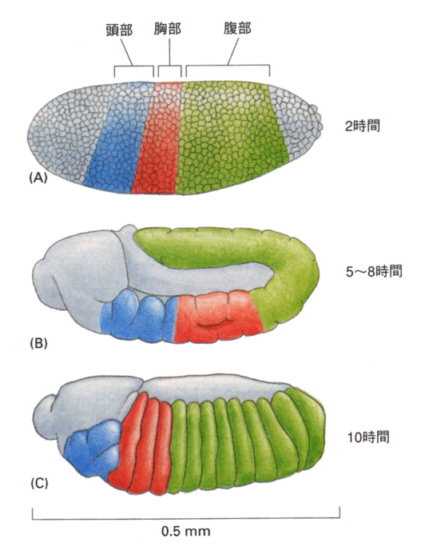
繰り返し構造であって、しかも運命が異なる体節がどのようにして分化するかについて、分子遺伝学的な手法を駆使してそのメカニズムが明らかになってきた。
2)母親由来のmRNA
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
http://www.ucalgary.ca/UofC/eduweb/virtualembryo/D_m_segment_I.html
最初の前後軸形成には母親の影響が大きい。卵が形成されるとき、生殖細胞が4回、分裂して16個の細胞になるが、お互いに細胞質の橋でつながっている。そのうちの1個が卵母細胞に、残りは哺育細胞になる。
哺育細胞は多くの成分を合成して卵母細胞に送り込む。卵母細胞を取り囲む濾胞細胞が末端シグナルを卵母細胞に送る。卵母細胞に送り込まれるのはmRNAで、bicoid mRNAが卵の前端に、nanos mRNAが後側に局在させる。これらのmRNAは、タンパク質でその場所に係留されていて、拡散しないようになっている。
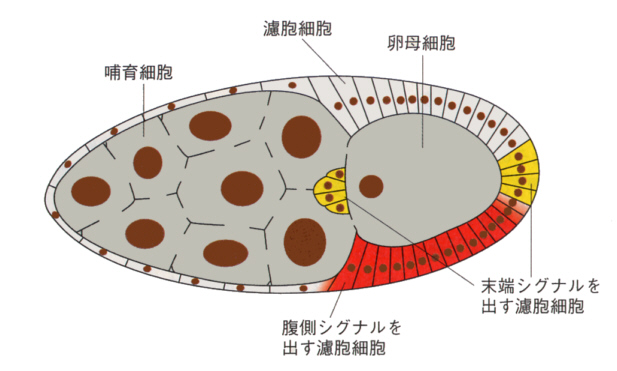
これらのmRNAが、発生の開始に伴って翻訳されてタンパク質が作られ、これらの濃度の勾配が、前後軸の形成に大きな影響を与える。
つまりbicoidタンパク質は前端が一番濃度が高く後ろに行くにしたがってしだいに薄くなるように、反対にnanosタンパク質は後端が一番濃くて前に行くにしたがって薄くなるような勾配ができる。
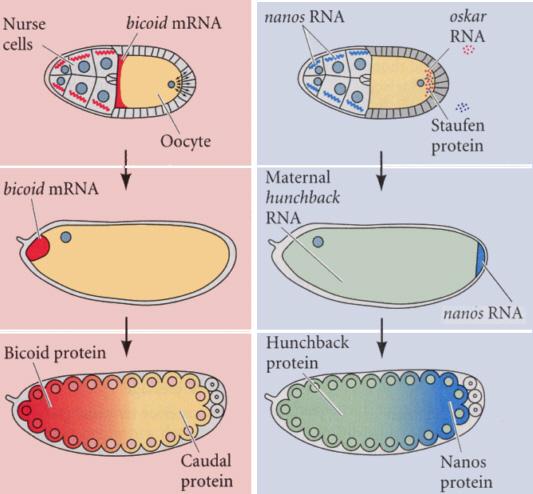
母親のbicoid遺伝子が突然変異で機能しなくなると、卵にmRNAが供給されず、したがってbicoidタンパク質の濃度勾配ができないので、この母親から生まれた卵からかえった胚には頭が無く、両端に尾部があるようになる。このように、胚の大まかな前後軸を形成に関係する遺伝子は、およそ20ほどが知られている。
bicoidタンパク質はどのように働いて頭部を形成するようにはたらくのだろうか。 bicoidタンパク質は転写調節タンパク質として、核の中に入り別のタンパク質の転写を活性化したり抑制したりする。次の遺伝子の産物であるタンパク質が、また次の遺伝子の転写を調節する、という連鎖が始まるのである。
3)分節遺伝子(segmentation genes)
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
http://www.ucalgary.ca/UofC/eduweb/virtualembryo/D_m_segment_II.html
大まかに前後軸(触れなかったが、実際は背腹軸も)が決まると、次にこれを基にして体節構造を形成する遺伝子が働く。胚に体節構造を作らせる遺伝子群を分節遺伝子と呼び、これには3つのグループがある。
1)ギャップ遺伝子(gap
genes)
2)ペアルール遺伝子(pair-rule genes)
3)セグメントポラリティー遺伝子(segment-polarity
genes)
である。これらの遺伝子は突然変異を詳細に分析することにより、明らかになった。
ギャップ遺伝子は、胚を大まかに区分けする働きがある。たとえば、Kruppel遺伝子は胸部第1節から腹部第5節までの8節の形成に関与している。hunchback遺伝子は下唇と胸部3節の形成に関与している。これらの突然変異体では、該当する体節を欠いた胚が生じる。
hunchback遺伝子は、母親由来のmRNAで卵全体に分布しているが、後部のnanosタンパク質などによって翻訳が抑制される。そのため、hunchbackタンパク質は、卵の前半だけに局限することになる(上の図参照)。
ギャップ遺伝子群のはたらきによって、卵は前、中、後部に分けられる。
ペアルール遺伝子は、ギャップ遺伝子群のはたらきによって大まかに分けられた各部域に体節構造を作り上げることに関与している。たとえば、ftz(fushi tarazu)の突然変異によって奇数番の体節を欠いた胚が生じ、eve(even-skipped)遺伝子の突然変異によっては偶数番の体節を欠いた胚が生じる。
下の図で、AからDはftzの発現がしだいに限局され、7本のバンドになるようすを示している。Eは青いeveタンパク質と茶色のftzタンパク質がバンド状に隣り合って発現しているようすを示す。こうして卵に体節構造ができあがる。
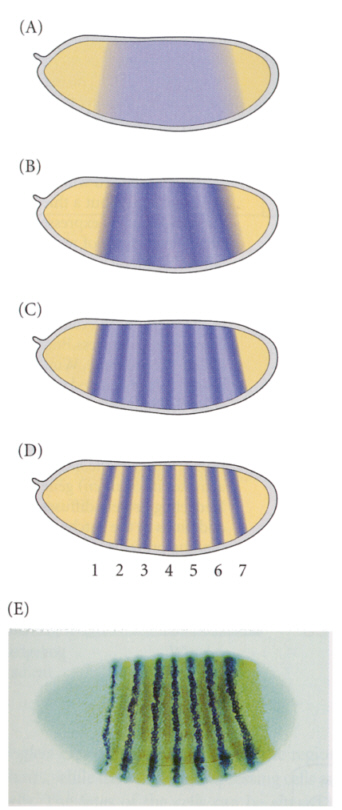
セグメントポラリティー遺伝子は、体節の数は正常だが、それぞれの体節の一部が欠失したり、ミラーイメージのものに置き代わったりする。たとえば、gooseberry突然変異体では、それぞれの体節の後半がすぐ直前の体節のミラーイメージになる。
それぞれの遺伝子のmRNAの発現場所や、発現したタンパク質の局在を調べてみると、1)から3)の順に発現して、母親由来のmRNAの局在によってギャップ遺伝子が発現しておおまかな区画割りがつくられ、ついでペアルール遺伝子が発現して体節をつくり、さらにセグメントポラリティー遺伝子が発現して、生じた体節構造を細かく規定して行くことが分る。
1)ホメオティック突然変異
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
http://www.ucalgary.ca/UofC/eduweb/virtualembryo/homeotics.html
ノーベル賞受賞講演(1955)pdfファイル
ホメオーシス(homeosis、相同異質形成)という現象が知られていた。たとえば、ザリガニの眼柄を除去すると普通は眼柄が再生するが、まれに触角が再生することがある。また、ショウジョウバエの突然変異で、触角の位置に肢が生えることがある。眼柄、触角、肢はいずれもそれぞれ本来生ずる体節に固有の付属構造である。相同の付属構造が場所を変えて生じる、これらの現象はホメオーシスと名付けられた(Bateson, 1894)。またホメオーシスを起こすような突然変異をホメオティック突然変異(homeotic mutation、相同異質形成突然変異)と呼ぶ。
2.で述べたように体節構造が決まると、それぞれの体節に固有な組織構造を作ることになるが、これを制御している遺伝子がホメオティック選択遺伝子(homeotic selector gene)である。これまで説明してきたように、特定の体節で発現した調節タンパク質が、その体節固有の構造を支配する遺伝子を活性化するのである。
ショウジョウバエでは、ホメオティック選択遺伝子は第3染色体上に2つある。一つはアンテナペディア遺伝子群(Antenapedia complex)で、もう一つは双胸遺伝子群(bithorax complex)である。群とあるように、それぞれ複数の遺伝子が近い位置に並んでいる(連鎖、linkage)。
体節に固有の構造を支配するので、この遺伝子の突然変異によって体の構造は大きく狂う。たとえば、アンテナペディア遺伝子の突然変異によって、下の写真のように、触角の位置に肢が生えてくる。
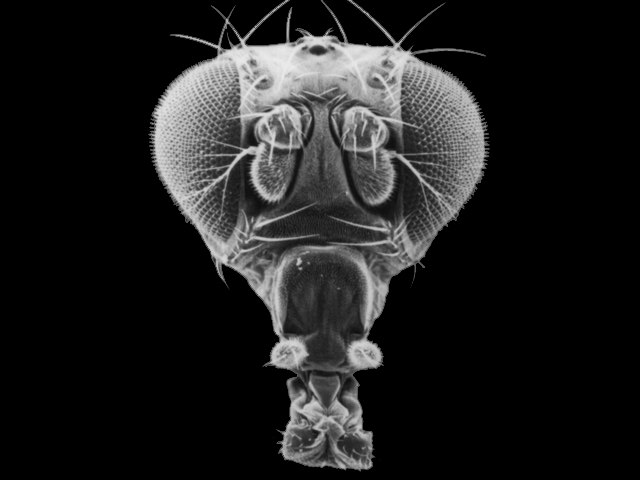

双胸遺伝子の突然変異によっては、胸部が二つできる。ショウジョウバエは双翅類というように普通は胸部第2節に1対の翅が生え、第3節には翅が変化した平均棍が生える。それがこの突然変異によって平均棍ではなく、ちゃんとした翅が生えた胸が二つになるのである。

2)ホメオボックス
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
http://www.biosci.ki.se/groups/tbu/homeo.html
これらのいくつもの遺伝子が解析され、その塩基配列が分ってくるとおもしろいことが分った。それはいずれの遺伝子にも180塩基の非常に相同性の高い部分が含まれていたのである。この部分はホメオボックス(homeobox)と呼ばれた。180塩基だからアミノ酸だと60個に相当する。タンパク質のこの部分をホメオドメイン(homeodomain)と呼ぶ。次の図はアンテナペディアホメオドメインが、DNAと結合しているようすである。
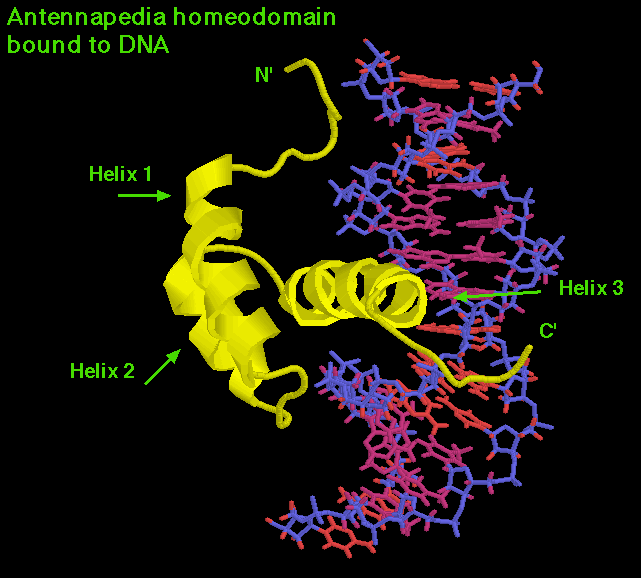
このように、ホメオドメインはDNAにはたらいて転写を調節するためのDNA結合部位(helix-turn-helix)だったのである。
次にホメオボックスを含むいくつかの遺伝子がコードするタンパク質を示した。赤い部分がホメオドメイン配列である。Antp, Ubxはショウジョウバエの遺伝子、Amphioxus Hox3はナメウジウオ(原索動物)、XenopusHOXA1はアフリカツメガエルから得られた遺伝子である。http://www.expasy.ch/cgi-bin/lists?hoxlist.txtよりSWISS-PROTへ跳び、作成した。
Drosophila Antp
1 11 21 31 41 51
1 MTMSTNNCES MTSYFTNSYM GADMHHGHYP GNGVTDLDAQ QMHHYSQNAN HQGNMPYPRF 60
61 PPYDRMPYYN GQGMDQQQQH QVYSRPDSPS SQVGGVMPQA QTNGQLGVPQ QQQQQQQQPS 120
121 QNQQQQQAQQ APQQLQQQLP QVTQQVTHPQ QQQQQPVVYA SCKLQAAVGG LGMVPEGGSP 180
181 PLVDQMSGHH MNAQMTLPHH MGHPQAQLGY TDVGVPDVTE VHQNHHNMGM YQQQSGVPPV 240
241 GAPPQGMMHQ GQGPPQMHQG HPGQHTPPSQ NPNSQSSGMP SPLYPWMRSQ FGKCQERKRG 300
301 RQTYTRYQTL ELEKEFHFNR YLTRRRRIEI AHALCLTERQ IKIWFQNRRM KWKKENKTKG 360
361 EPGSGGEGDE ITPPNSPQ
Drosophila Ubx
1 11 21 31 41 51
1 MNSYFEQASG FYGHPHQATG MAMGSGGHHD QTASAAAAAY RGFPLSLGMS PYANHHLQRT 60
61 TQDSPYDASI TAACNKIYGD GAGAYKQDCL NIKADAVNGY KDIWNTGGSN GGGGGGGGGG 120
121 GGGAGGTGGA GNANGGNAAN ANGQNNPAGG MPVRPSACTP DSRVGGYLDT SGGSPVSHRG 180
181 GSAGGNVSVS GGNGNAGGVQ SGVGVAGAGT AWNANCTISG AAAQTAAASS LHQASNHTFY 240
241 PWMAIAGECP EDPTKSKIRS DLTQYGGIST DMGKRYSESL AGSLLPDWLG TNGLRRRGRQ 300
301 TYTRYQTLEL EKEFHTNHYL TRRRRIEMAH ALCLTERQIK IWFQNRRMKL KKEIQAIKEL 360
361 NEQEKQAQAQ KAAAAAAAAA AVQGGHLDQ
Amphyoxus Hox3
1 11 21 31 41 51
1 MQKSPYYETT SQQLYNGYSY SNGERFGYEG SGGGGGGGSY GAEVTQDFGP SCSVRKPGST 60
61 TGGGTGNVSP TCMKTNPADN NHGSAPQSNA SILANTKIYP WMKESRQNSK QRQQPNLSVG 120
121 TTEPGESPGL GGAAGKRART AYTSAQLVEL EKEFHFNRYL CRPRRVEMAA MLNLTERQIK 180
181 IWFQNRRMKY KKEQKVKGGG SGGGSGGMNS PSPPATTTPP GINPGPLPHP TTQPSLSQSN 240
241 NMSNHMSMMG SLQQTQPAYS QYPPPHLNHS LPHQAPPHSV GLTMSGPVSP QCYQSNHSPC 300
301 PPSSAPHPVP MGNHVPHPRQ GPPHMTNGLP ASPLEVVSQR QYTPPAPGPS PTGPGPGHHG 360
361 LTNSYPECPP HHTELPHHQY STPMSVVNMN YCVNVQTQGN RLNSAPKLTH L
Chick HOXA2
1 11 21 31 41 51
1 MNFEFEREIG FINSQPSLAE CLTSFPPVGD TFQSSSIKNS TLSHSTLIPP PFEQTIPSLN 60
61 PGGHPRHSGG GRPKASPRAR SGSPGPAGAP PPPEYPWMKE KKASKRSSLP PASASASAAG 120
121 PACLSHKDPL EIPDSGSGGS RRLRTAYTNT QLLELEKEFH FNKYLCRPRR VEIAALLDLT 180
181 ERQVKVWFQN RRMKHKRQTQ CKENQNGEGK FKGSEDPEKA AEDDEEEKAL FEQALGTVSG 240
241 ALLEREGYAF QQNALSQQQA QNAHNGESQS FPVSPLTSNE KNLKHFQHQS PTVQNCLSTM 300
301 AQNCAAGLNN DSPEALEVPS LQDFNVFSTD SCLQLSDAVS PSLPGSLDSP VDISADSFDF 360
361 FTDTLTTIDL QHLNY
Human HOXA1
1 11 21 31 41 51
1 MDNARMNSFL EYPILSSGDS GTCSARAYPS DHRITTFQSC AVSANSCGGD DRFLVGRGVQ 60
61 IGSPHHHHHH HHHHPQPATY QTSGNLGVSY SHSSCGPSYG SQNFSAPYSP YALNQEADVS 120
121 GGYPQCAPAV YSGNLSSPMV QHHHHHQGYA GGAVGSPQYI HHSYGQEHQS LALATYNNSL 180
181 SPLHASHQEA CRSPASETSS PAQTFDWMKV KRNPPKTGKV GEYGYLGQPN AVRTNFTTKQ 240
241 LTELEKEFHF NKYLTRARRV EIAASLQLNE TQVKIWFQNR RMKQKKREKE GLLPISPATP 300
301 PGNDEKAEES SEKSSSSPCV PSPGSSTSDT LTTSH
Mouse HOXA1
1 11 21 31 41 51
1 MNSFLEYPIL GSGDSGTCSA RAYPSDHGIT TFQSCAVSAN SCGGDDRFLV GRGVQISSPH 60
61 HHHHHHHHHH PQTATYQTSG NLGISYSHSS CGPSYGAQNF SAPYGPYGLN QEADVSGGYP 120
121 PCAPAVYSGN LSTPMVQHHH HHQGYAGGTV GSPQYIHHSY GQEQQTLALA TYNNSLSPLH 180
181 ASHQEACRSP ASETSSPAQT FDWMKVKRNP PKTGKVGEYG YVGQPNAVRT NFTTKQLTEL 240
241 EKEFHFNKYL TRARRVEIAA SLQLNETQVK IWFQNRRMKQ KKREKEGLLP ISPATPPGSD 300
301 EKTEESSEKS SPSPSAPSPA SSTSDTLTTS H
Xenopus HOXA1
1 11 21 31 41 51
1 AYAHSSCGSN YGMQNFSPGY SHFPIHQETE VSSGFPQSVY SGNIASSVVQ HQQHQSYIEG 60
61 SAHYIHHSYG PEQNLSVANY NNNVASLHIS QREVCHSPSS ETSPGPTQTF DWMKVKRNPP 120
121 KTGKAGEYGY AGQPNTARTN FTTKQLTELE KEFHFNKYLT RARRVEIAAA LQLNETQVKI 180
181 WFQNRRMKQK KREKEGLLPI SPSTSTGSDE KSEELSEKSN SSPCAPSPAS STSDHLSASG
Zebrafish HOXA1
1 11 21 31 41 51
1 MNSYLDYTIY NRGSNTYSSK VGCFPVEQEY LPSACASTNS YIPEGRPVGG NTFTSAPHET 60
61 HGTSYAQIQS QAFHLNVDMG KTGHSNFCKQ TRPPHSDYGH QHVLTQADDH MRLQSPGFSV 120
121 VNMGANIGTY SESNCRPGSV SASHYQSYAY GEPEPHGCGS FSKYQVSPDS DSDSKTNIKQ 180
181 APTFDWMKVK RNPPKTVKVA EYGIHGQQNI IRTNFTTKQL TELEKEFHFN KYLTRARRVE 240
241 VAATLELNET QVKIWFQNRR MKQKKREKEG TAPVIKRVTL CSSGQNRRPL KLAHRLVPSP 300
301 TSDSSTAI
上の配列から、ホメオドメイン部分だけを抜き出した比較したのが次の図である。赤いアミノ酸は一番上に示したAntpの配列と異なる場合を示している。ショウジョウバエからヒトまで共通した部分が存在することがわかるだろう。
Antp RKRGRQTYTR YQTLELEKEF HFNRYLTRRR RIEIAHALCL TERQIKIWFQ NRRMKWKKEN Ubx RRRGRQTYTR YQTLELEKEF HTNHYLTRRR RIEMAHALCL TERQIKIWFQ NRRMKLKKEI Am HOX3 GKRARTAYTS AQLVELEKEF HFNRYLCRPR RVEMAAMLNL TERQIKIWFQ NRRMKYKKEQ ChickHOXA2 SRRLRTAYTN TQLLELEKEF HFNKYLCRPR RVEIAALLDL TERQVKVWFQ NRRMKHKRQT HumanHOXA1 PNAVRTNFTT KQLTELEKEF HFNKYLTRAR RVEIAASLQL NETQVKIWFQ NRRMKQKKRE
さらに上の配列から、脊椎動物のHOXA1のホメオドメイン部分だけを抜き出した比較したのが次の図である。一番上のヒトのホメオドメインと異なるアミノ酸は緑で示してある。脊椎動物の中では、ホメオドメインがほとんど同じだということがわかる。
HumanHOXA1 PNAVRTNFTT KQLTELEKEF HFNKYLTRAR RVEIAASLQL NETQVKIWFQ NRRMKQKKRE MouseHOXA1 PNAVRTNFTT KQLTELEKEF HFNKYLTRAR RVEIAASLQL NETQVKIWFQ NRRMKQKKRE XenopusHOXA1 PNTARTNFTT KQLTELEKEF HFNKYLTRAR RVEIAAALQL NETQVKIWFQ NRRMKQKKRE ZebrafishHOXA1 QNIIRTNFTT KQLTELEKEF HFNKYLTRAR RVEVAATLEL NETQVKIWFQ NRRMKQKKRE
このように、ホメオドメインを含むこれらのタンパク質は、DNAに結合して転写を調節する重要な分子であることが分った。現在では、ホメオティック遺伝子(あるいはホメオタンパク質)という言葉には、最初の相同異質形成(ホメオーシス)という意味は薄れ、ホメオボックスを含む遺伝子(あるいはホメオドメインを含むタンパク質)という意味合いが強くなっている。
ショウジョウバエで見つかったこの配列は、上の例にあるように生物に共通した配列である。これは、ホメオボックスから得たcDNAをプローブとして、他の生物のゲノムをサザンブロット法で解析することにより明らかにされた。
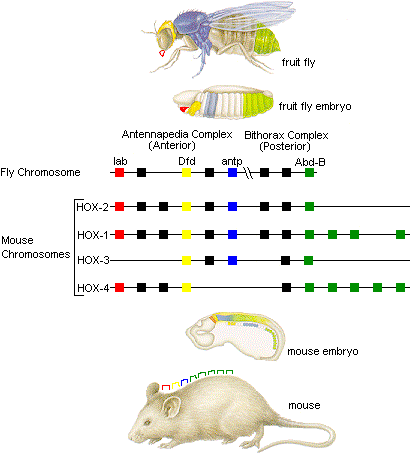
このように、ホメオティック遺伝子は、長い進化の歴史で保存されてきたDNAの領域であることから、重要な働きを持った部分であることが推測された。ショウジョウバエでも哺乳類でも、この遺伝子は発生の過程で身体の構成を順序立てる重要な働きをしていることが分かっている。
脊椎動物の場合は、発生する細胞の運命は、これまで述べた昆虫の場合ほど初めから厳密に決まっていない。始原生殖細胞の場合は例外的に早い時期に決まるが、その他の細胞の場合は、他の細胞からの影響で決まってくる。したがって発生がある程度進んだ場合でも、例えば胚を半分にしても両方から完全な個体が発生する場合がある。これは細胞の運命をある程度、後になっても変更できるためである。
このような卵を調節卵と呼ぶことはすでに述べた。それでは細胞の運命はどのように決まるのだろうか。有名なシュペーマンの二次胚の実験が示すように、他の細胞からの誘導(induction)の結果、細胞の運命は徐々に決まってくる。
そこで重要になるのが、細胞間の位置関係である。嚢胚形成(gastrulation)が最初の細胞構築にとって重要であることがこれで理解できる。胞胚(blastula)になることで、卵細胞質の分配が行われ、胞胚の各細胞の運命がある程度決まる。ついでこれが嚢胚(gastrula)になり、二重になることで裏と表の関係ができる。裏側の細胞がすぐ上に来た表の細胞に働きかけて、その運命を決定する。
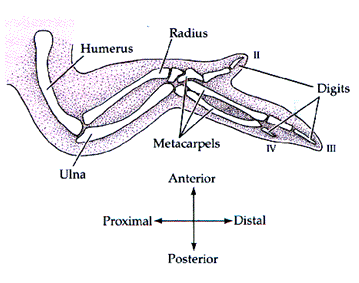
脊椎動物でも、こうして誘導と決定の連鎖が進んで行くことになる。ここではニワトリをつかった肢の発生を、ごく簡単に見てみよう。肢には上に述べた軸がすべて必要だからである。
ニワトリの前肢(後に翼となる)は次の図のような構造をしている。指は2本欠損している。
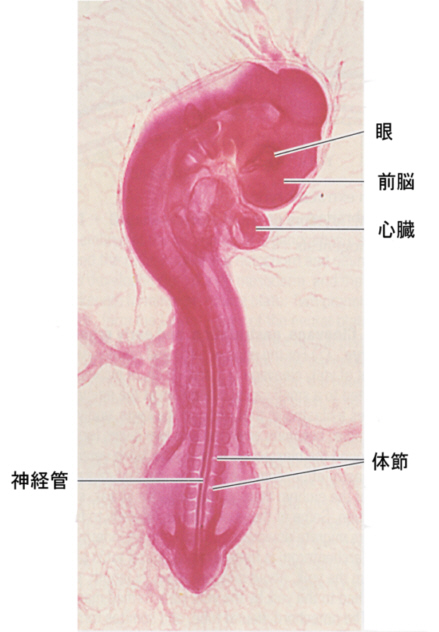
下の方に載せてある、鳥類の初期発生の最後の図から少し発生が進んだ頃、身体の両側、心臓の位置よりかなり下の方と後端に近い部分に膨らみが現れてくる。これが将来、肢になる肢芽(limb bud)である。
この肢芽は、移動してきた中胚葉である間充織細胞群が分泌する、FGF(fibroblast growth factor、繊維芽細胞増殖因子)の一つによって誘導される。別な転写調節因子が間充織にはたらいて、前肢になるか後肢になるかの方向づけをおこなっている。こうして間充織と表皮でできた肢芽ができ、その先端の表皮は厚くなってAER(apical ectodermal ridge、外胚葉性頂堤)を形成する。
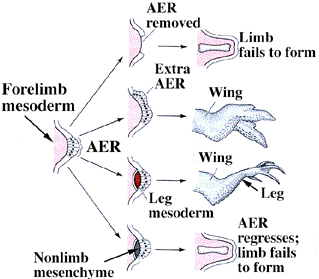
AERは別のFGFを分泌して肢芽の成長を促すとともに、肢芽の後部の間充織にはたらきかけてZPA(zone of polarizing activity、極性化活性帯)をつくる。この部分を別の個体の肢芽の前端に移植すると、真ん中から鏡像関係を持った指が生じ、この部位が肢芽の前後の極性を作っていることがわかる。ZPAからはshh(sonic hedgehog)という遺伝子からタンパク質ができている。
これとは別(Wnt7a)という情報がもう一つの軸、背腹軸を作っている。
さらに、これらの過程と平行してホメオボックス遺伝子が順番に発現して、肩甲骨から肢指までの骨を正しい順番に形成していく。
これ以上は複雑になるのでここでは省略する。
終わりは次の始まりである。この講義はこれで終わるが、次の道がさらに先へと伸びている。細胞生物学のテキストの最後ではFollow the yellow brick road!! と書いて、映画「オズの魔法使い」のドロシーと3人(?)の例を出したが、黄色い道は一直線にエメラルドシティーへつながっているのではない。途中悪い魔法使いの妨害もあるだろう。けれども基本的には学ぶことは楽しいことだ。
3学期になると、いよいよこれまでの学んできたことを基礎として、専門過程の学習が始まる。人間の病態を知るには、まず正常な形態、生理を学ぶ必要がある。そのために、どのように細胞が組織化され器官や器官系が作られているか、個体を取り囲む環境の情報をどのように受け取り処理しているか、細胞間の調整をどのような信号分子でどのように行っているか、外敵の攻撃から個体をどのように防衛しているか、などをさらに詳しく学習することになる。そのための基礎としてこれまで学習したことが役に立てば幸いである。
何故だろう、どうなっているのかしら、という問題意識を常に持って、自分で調べてみるという気持ちを忘れずに、生命科学探求の道を進んで欲しい。
1)鳥類の場合
器の中に割って中身を出したニワトリの卵を想像して欲しい。卵黄が多いので、細胞質は黄身の上にほんのわずかに白く見える部分に限られる。卵割はこの部分で進行する。
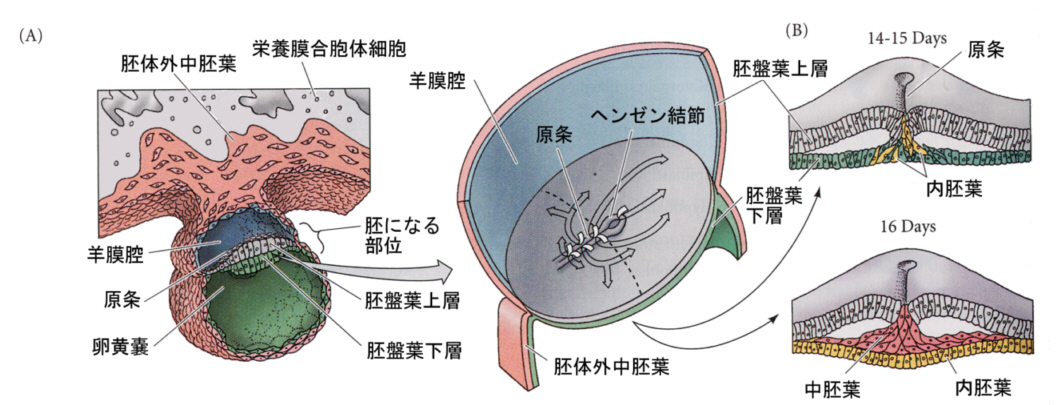
卵割が進むと割球の数が増え、胚盤を形成する。この薄い胚盤が二層になり、さらに原条と言う切れ込みが前後にでき、ここから陥入が始まる。
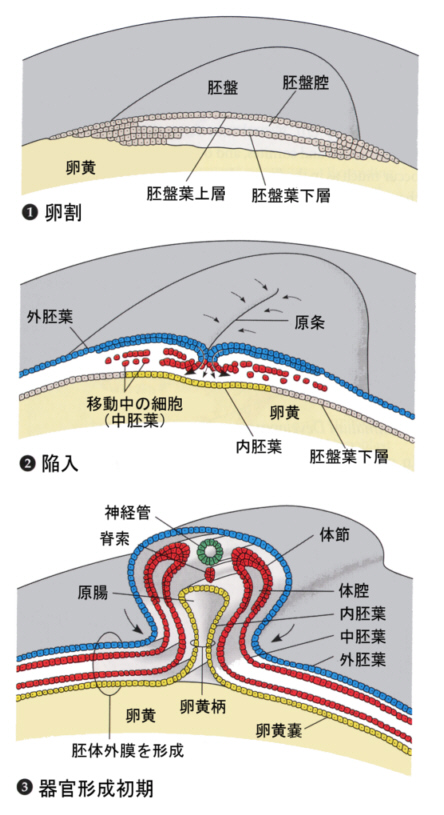
三胚葉ができた後は、大きな卵黄嚢がある以外は、カエルの発生と本質的には似ている。
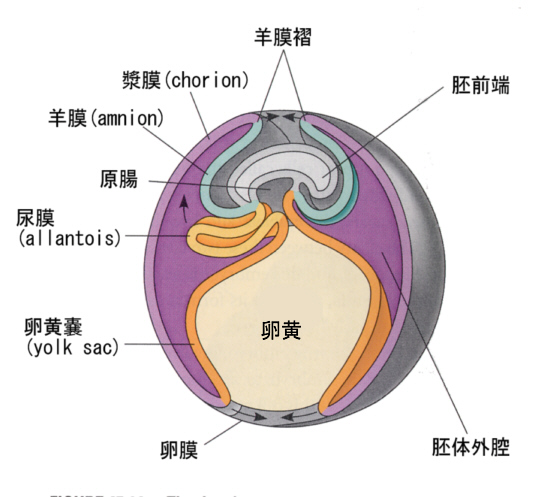
その後、胚体外に漿膜(chorion)、羊膜(amnion)、尿膜(allantois)ができ、羊膜は胚を包むようになり、以降胚は羊膜腔の中で発生が進む。下の図は胚を実際よりも模式的に大きく描いている。
2)哺乳類の場合
哺乳類では、体内で受精して子宮内膜へ着床し、母体内で発生が進む(胎生)ので、これまで述べてきた発生のようすとは少し異なった経過をたどる。
排卵された卵が輸卵管を下る過程で、交尾によって膣内に送り込まれた精子が膣から子宮への入り口を突破して輸卵管へ入り、ここで卵と受精する。受精卵は卵割をしながら輸卵管を下っていく。
8細胞期まではほぼ等割していき、各割球は球形でおたがいにゆるい結合をしているが、8細胞期の終わりに急激に割球の形が変わり、各割球がお互いに密接するようになる。この現象をコンパクションと呼んでいる。これは各割球の間にタイトジャンクションと接着タンパク質(E-カドヘリン)による接着ができるからである。この過程にはPKCが関与していると考えられている!
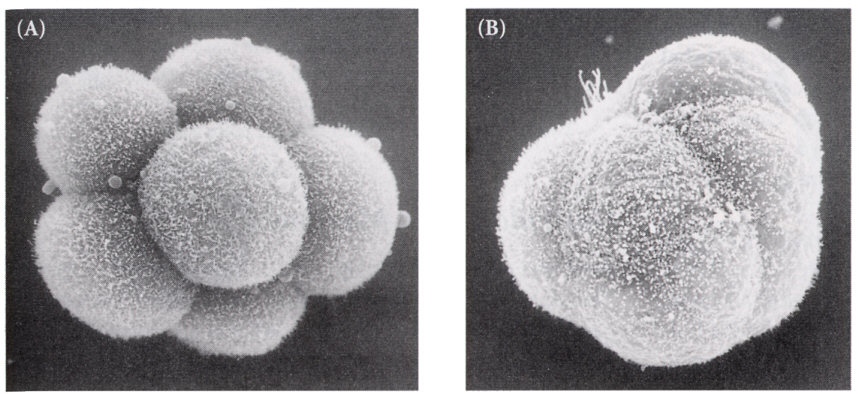
次の卵割により16細胞期になったときに、内側になった割球と外側の割球に別れ、外側の割球はタイトジャンクションと接着タンパクで結合し、内側の細胞はギャップジャンクションで結合するようになる。
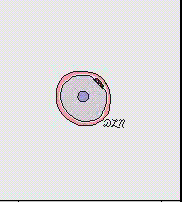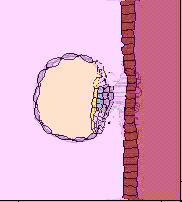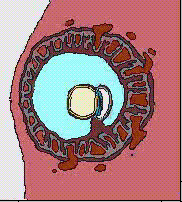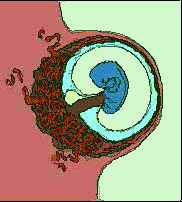
32細胞期になると、外側の細胞は水を吸い込んで一層の細胞層になり、内側の細胞塊が上部にへばりついた形となる。こうして外側の細胞層である栄養芽層(trophoblast)と、内部の細胞群である内部細胞塊(inner cell mass)に分かれる。栄養芽層は後に胎盤の形成に参加し、内部細胞塊のみが胚として発生をすることになる。この時期は他の動物の胞胚に該当するが、胚盤胞(blastcyst)と呼び、この時期に着床する。
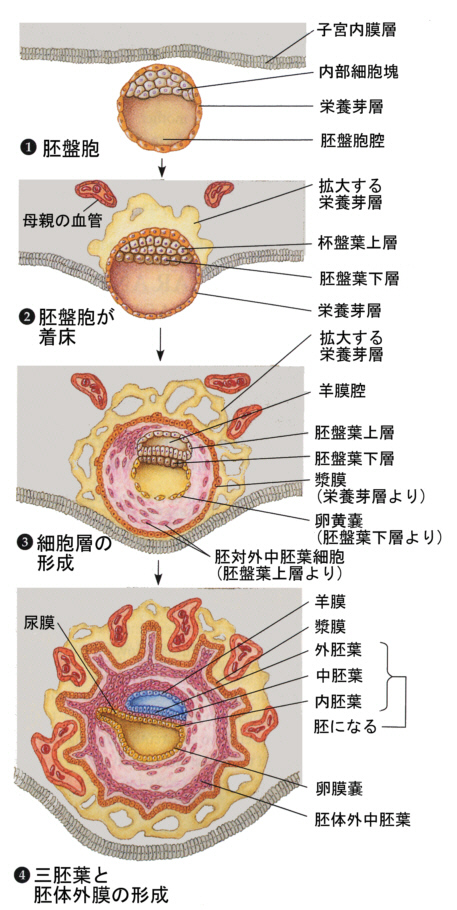
子宮内に着床してできた円盤状の胚葉が、ちょうど鳥類の胚盤に該当すると考えればよい。ここに原条ができ、陥入がおこり、胚体外膜は胎盤を形成することになる。
ヒトの発生をさらに調べたい人へ(このページへ戻るときはブラウザーの戻るを選んでください)
http://embryo.soad.umich.edu/index.html
![]() この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
 ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。
ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。