1.神経系 2.神経細胞(ニューロン、neuron) 3.膜電位 4.活動電位 5.活動電位の伝導 関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください) Neuroscience for kids Lecture notes for Behavioral Neurobiology of Concorde University On-Line Biology Book: The Nervous System 更新日:2001年8月31日
1)いろいろな神経系
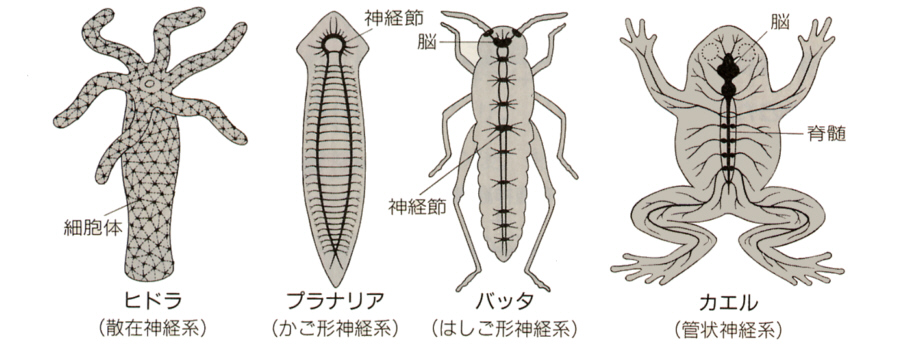
神経系は原生動物と海綿動物を除く動物にみられ、動物の種類(進化の程度)によって構造が異なっている。ヒドラやクラゲなどの神経系は、神経細胞が網の目のように連絡した神経網を形成している。このような神経系を散在神経系という。
プラナリアなどでは頭部に神経細胞体が集まって神経節を作るようになる。ミミズやゴカイなどの環形動物やエビやバッタなどの節足動物では、体節ごとに神経節が1対づつあり、これらが神経繊維でつながったはしご形神経系を形成する。頭部の神経節は発達して脳と言われるようになり、頭部集中化がすすむ。
脊椎動物では、すべての神経細胞は神経管に由来する。神経管を構成する細胞が分化して、いろいろな神経細胞を作り出す。
2)中枢神経系と末梢神経系
神経管は脳や脊髄になり、これらを総称して中枢神経系(Central Nervous System)と呼ぶ。中枢神経系の外にある神経細胞は、神経冠から生じるが、これももとをただせば神経管から分かれたものである。中枢神経系の外にある神経を末梢神経系(peripheral nervous system)と呼ぶ。
末梢神経系はさらに体性神経(somatic nervous system)と自律神経(autonomic nervous system)に分けられる。
体性神経は感覚器からの感覚情報を中枢に伝える求心性神経と、中枢の司令を筋肉や腺に伝える遠心性神経からなる。
3)自律神経系
自律神経系は大部分が中枢からの遠心性神経で、意志によっては制御できない。さらに交感神経系(sympathetic nervous system)と副交感神経系(parasynpathetic nervous sytem)とに分けられる。自律神経の支配を受ける器官はふつう、交感神経と副交感神経の両方が分布していて、多くの場合、一方がそのはたらきを促進すれば他方は抑制するというように、拮抗している。
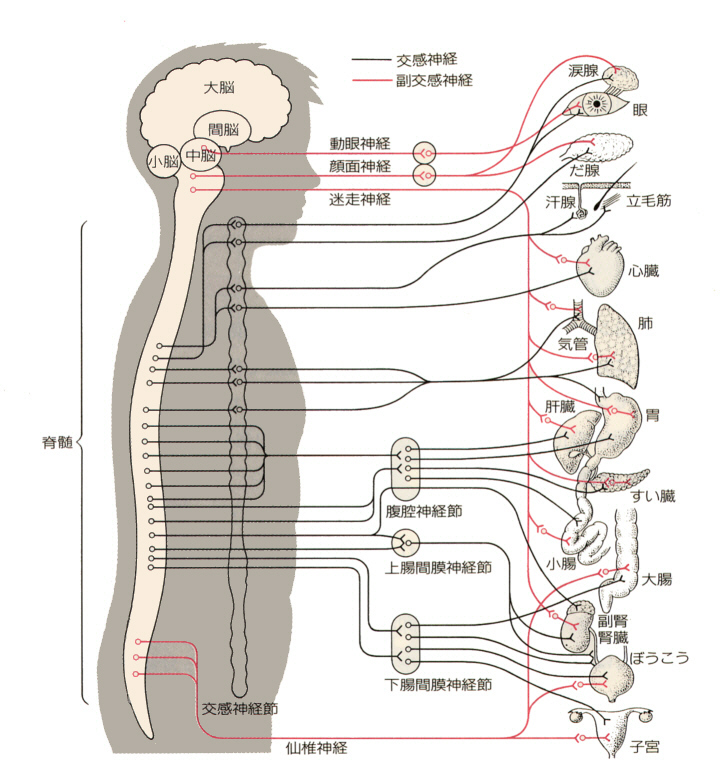
交感神経は、脊髄から出て交感神経節や腹腔神経節で神経を乗り換え、ここから心臓、肺、消化管などの器官に分布している。副交感神経は、中脳から出る動眼神経、延髄から出る顔面神経や迷走神経、脊髄の末端から出る仙椎神経などがあり、分布する各器官のそばの神経節で神経を乗り換えてそれぞれの器官に分布する。
中枢から神経節までの神経繊維を節前繊維(pregannglionic fiber)、神経節から先の神経繊維を節後繊維(postsynaptic fiber)と言う。
自律神経系についてさらに学びたい人へ
http://gwis2.circ.gwu.edu/~atkins/Neuroweb/autonomic.html
神経系についてさらに学びたい人へ
http://thalamus.wustl.edu/course/
1)神経細胞とグリア細胞
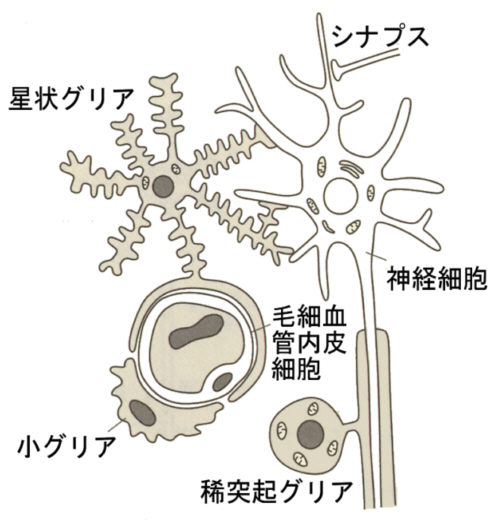
神経系を構成する細胞は、神経細胞とグリア細胞である。グリア細胞は神経系の情報伝達の機能とは関係せず、神経細胞を側面から支える役割をする。
グリア細胞にはグリア細胞のグリアとはギリシャ語で膠(にかわ)と言う意味で、神経細胞にぴったりと寄り添っているのでこの名がある。グリア細胞には、神経細胞と血管の間にあって、神経細胞へ栄養を供給している、突起の多い星形をした星状グリア細胞(アストロサイト、astroglia)と、突起は少なく、軸索に巻き付いて髄鞘を形成する稀突起グリア細胞(oligodendroglia)がある。この他に血管近くにあるミクログリア(microglia)があり、脳におけるマクロファージであると考えられているが、まだ不明な点も多い。
神経細胞は実にさまざまな形態を取る。
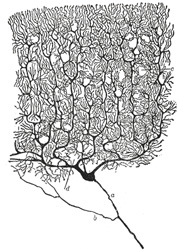
大脳皮質の錐体細胞 小脳のプルキンエ細胞
神経細胞を一つの単位と考えてニューロン(neuron)と呼ぶ。
2)運動ニューロンの形態
ここではとりあえず典型的な運動ニューロンの形を考えてみよう。運動ニューロンは次の図のような形態をしている。
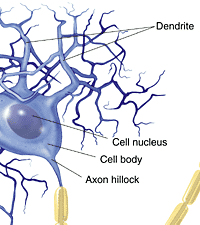


http://cw.prenhall.com/bookbind/pubbooks/morris2/chapter2/custom1/deluxe-content.htmlより
ニューロンと言えども細胞の一種なので、基本的な作りはこれまでに学んだ細胞の作りと同じである。大きく異なるのは、2種の顕著な突起を有することである。一つは枝分かれした樹状突起(dendrite)、もう一つは1本の長い軸索(axon、neurite)である。核を含む細胞体の部分(cell body、soma)には、粗面小胞体とゴルジ体がよく発達している。
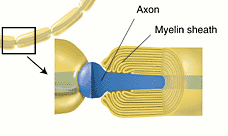
軸索が細胞体から出るところは、少し太くなっていて、軸索丘(axon hillock)とか軸索起始部(initial segment)と呼ばれる。運動ニューロンの軸索はシュワン細胞が巻き付いてミエリン鞘(myelin sheath)を形成している。ミエリン鞘とミエリン鞘の間には切れ目があり、これをランビエ絞輪(Ranvier's node)と呼ぶ。
3)ニューロンの機能的分化
ニューロンはそれぞれの部域で機能的な役割の分担をしている。樹状突起部は、他からの情報を受け取る場所である。樹状突起(と細胞体)には多数のシナプスが形成されている。情報はここから入力される。細胞体部はおもに樹状突起で受けた入力を統合する場所となる。
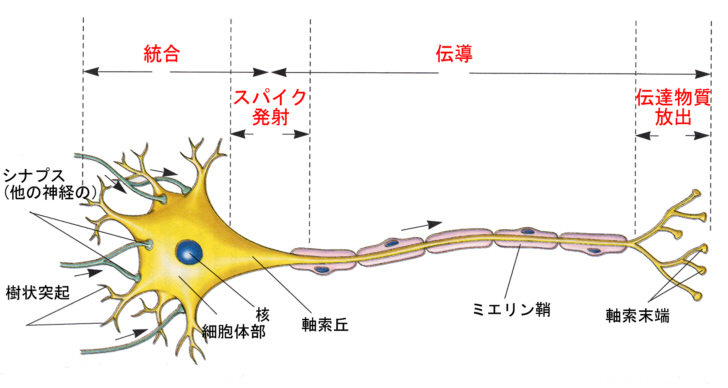
統合された情報は軸索丘に伝えられ、ここでインパルスが発射される。つまり、ある一瞬にインパルスを出すか出さないかというように、デジタル化される。
生じたインパルスは、軸索を伝導(conduction)していく。軸索末端である終板までインパルスが達すると、神経伝達物質(neurotransmitter)の放出が起こり、効果器である筋肉が収縮する。
終板というのは運動ニューロンと筋肉の特殊化したシナプスであるが、中枢神経系内でも、ニューロンと次のニューロンの樹状突起あるいは細胞体との間にシナプスが形成され、神経伝達物質によって情報が伝達される。一般的なシナプスは次のような構造をしている。
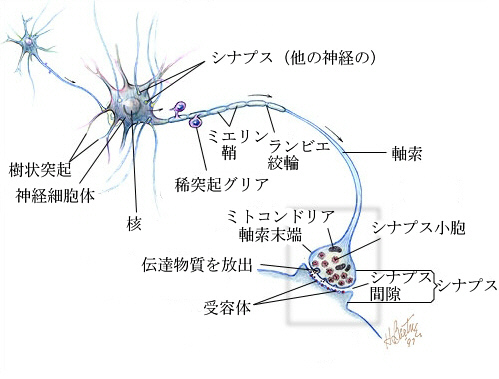
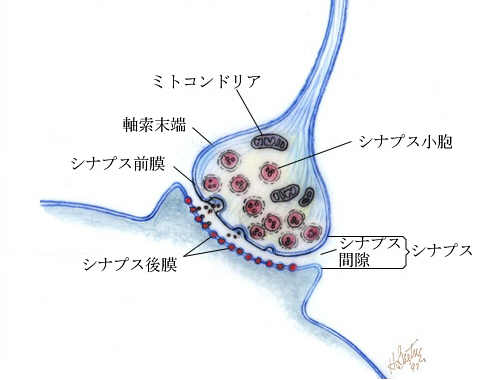
シナプスは、シナプス前膜(presynaptic membrane)、シナプス間隙(synaptic cleft)、シナプス後膜(postsynaptic membrane)で構成されている。
こうして、情報が樹状突起から軸索末端へと伝えら、次の要素(筋肉や腺などの効果器あるいは次のニューロン)へ伝えられいく。信号は電気的→化学的→電気的と変換されていることになる。
それではどうしてこのようなことが起こるのか。これを解く鍵は、細胞膜に埋め込まれた電位依存性のイオンチャンネルなどの膜タンパク質である。
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
Neuroscience for kids
- action potential
神経や筋肉が、情報を伝えたり機能を発揮できるのは、巧みなイオンの移動機構により一瞬に近い早さで電位変化を起こすことができるためである。このような性質を持った膜を興奮性膜(excitable membrane)と言う。それではどのようなメカニズムによって興奮するのであろうか。
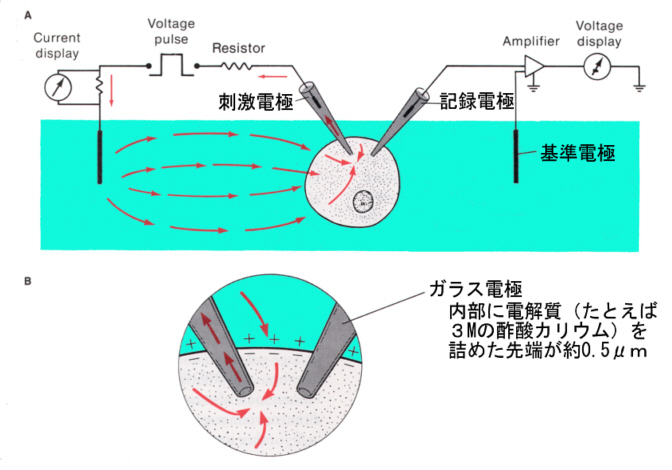
興奮性膜を持った細胞(すなわち神経細胞や筋肉の細胞)を生理食塩水に入れ、生理食塩水中に不関電極(reference electrode)をおき、ガラス電極と不関電極の間の電位を測りながら電極を細胞に近付けて行くと、はじめは電位差は0であるが、細胞に入った瞬間に-70から-90mv(細胞内が負)の電位差が計測できる。以後、細胞が活動しない限り、この電位差は保たれる。
この電位差を静止電位(resting potential)という。この静止電位はどうして発生するのだろうか。
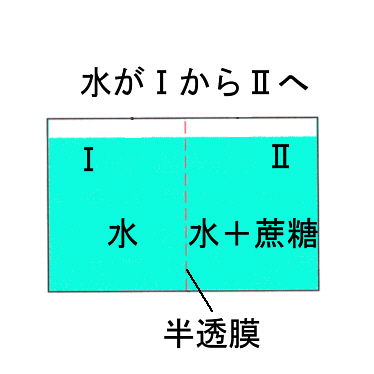
①浸透圧(Osmosis)
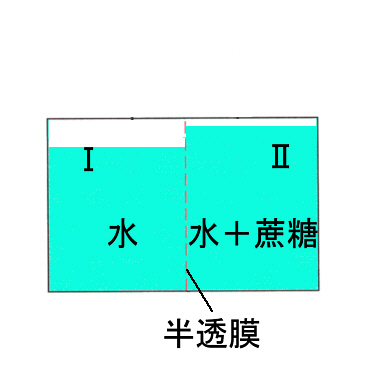
次の図のように、真中を半透膜で仕切った片方(Ⅰ)に水を入れ、反対側(Ⅱ)に蔗糖溶液を入れると、やがて水の入っていた方が低く蔗糖液の方が高くなり安定する。この状態では、Ⅰ内の液が膜に働いている浸透圧(osmotic pressure)と、ⅠとⅡの差に相当する水圧(hydrostatic pressure)で膜に働く力が釣りあっている。
②平衡電位(equilibrium potential)
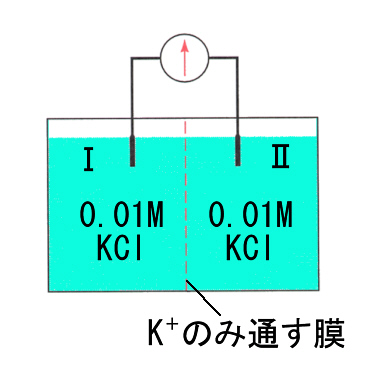
①と同様に、K+のみを通す半透膜を仮定し、半透膜で仕切った両側に0.01M KClの水溶液を入れると、個々のK+ を見れば移動する場合があるが、全体としては移動はなく、電気的に中性で、左側の槽(Ⅰ)と右側の槽(Ⅱ)の間には電位差はない。
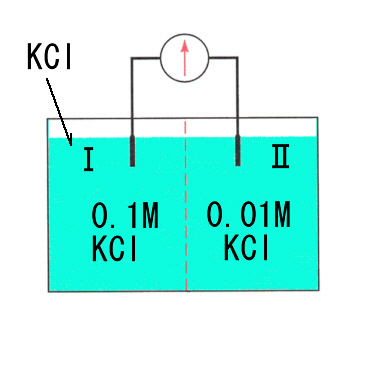
いま(Ⅰ)にKClを加え濃度を0.1Mにすると、 K+の濃度差ができるので K+ は濃度勾配にしたがって(Ⅱ)へ移動しようとする。
しかし膜の近辺では、 K+が移動したことによる(Ⅰ)側の負によって生じるK+を(Ⅰ)に引き戻そうとする力(electromotive force)と、濃度勾配による(Ⅰ)→(Ⅱ)の移動(diffusion)が釣りあって、すぐに定常状態になる。
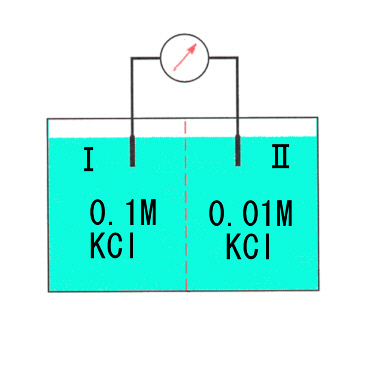
このとき、(Ⅰ)と(Ⅱ)の間には次の式で決まる電位差が生じる(平衡電位、equilibrium potential)。
![]()
ここで[X]はモル濃度、Rはガス常数、Tは絶対温度、FはFarady常数、zはイオンの価数である。
[ K+]は一価だから上の式を常用対数になおすと
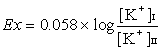
ボルト(18度Cの場合)になる。上の例では
![]()
=58mV になる。
このような関係が成り立つのは、 K+のみを通す半透膜を仮定したからである。もしも仕切り膜が、鉄板やプラスチック板で、K+を通さなければ、このようなことは起こらない。このようにK+のみを通す半透膜の両側にK+のイオンの濃度差が存在すると平衡電位が成立し、一種の電池ができることになる。イオンが電荷を運ぶキャリアーだが、実際にはイオンの移動は濃度が変化する程は起こらない。
上の例では人が2槽間のK+の濃度差を作ったが、細胞ではこのような濃度差はATPのエネルギーを使ってポンプ(Na+-K+ ATPase)が作り出している。
実際の細胞に見られるイオンの偏りは次の表のようである。
| 細胞内(mM) | 細胞外(mM) | |||||
| Na | K | Cl | Na | K | Cl | |
| イカgiant axon | 49 | 410 | 40 | 440 | 22 | 560 |
| カニ神経 | 52 | 410 | 26 | 510 | 12 | 540 |
| カエル縫工筋 | 15 | 125 | 1.5 | 110 | 2.6 | 77 |
特に注目しなければならないのはNa+とK+で
Na+:細胞外に多く、細胞内に少ない
K+ :細胞外に少なく、細胞内に多い
興奮性膜を持った細胞が興奮していない(静止時)ときは、膜はK+に対する透過性のみ存在し、その他のイオンに対する透過性は極めて小さい(K+に対するチャンネルのみが開いている)。そうすると静止電位は次の式から計算できることになる(上の表のK+ の濃度を用いて計算してみよ)。
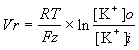
![]()
(ただし18度Cで)
ガラスの先を細くして塩化カリウムを詰めたガラス電極を細胞内に刺入して、実際に静止時の電位を測定してみると、計算値とほぼ一致する。
神経が使うシグナルはただ一つ、活動電位(action potential)であり、活動電位を発生して、これを次の神経(あるいは筋肉)へ軸索を通してつたえる。
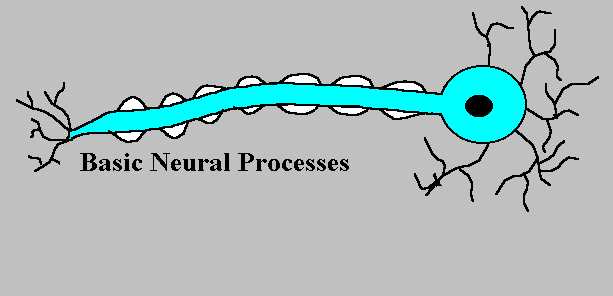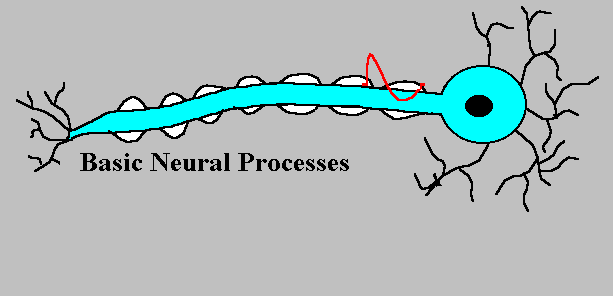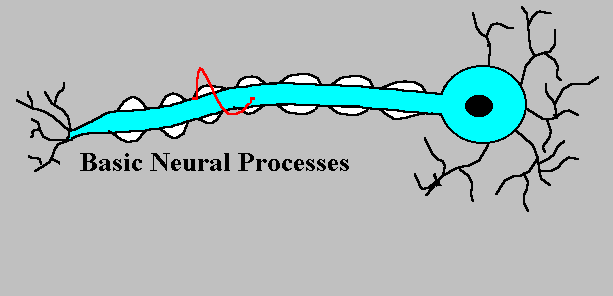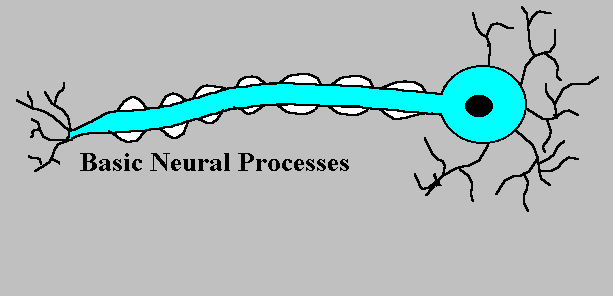
活動電位は、大きな、きわめて短い電位の変化である。活動電位はいったん発生すると、どのような方法で刺激されたかに関係なく、同一の神経では同じ経過をたどり、したがって「全か無か(all-or-none)の法則」に従うと言う。
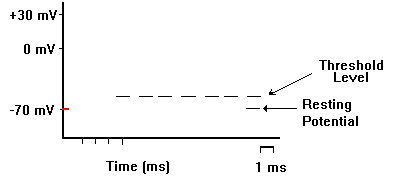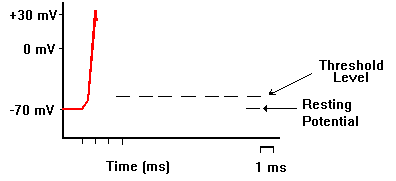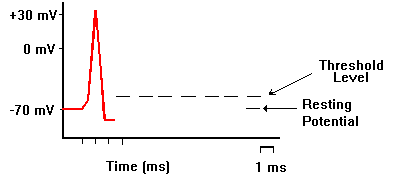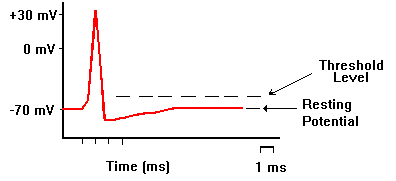
記録用のガラス電極を刺入して、別の刺激電極で刺激すると、活動電位を生じさせ、記録することができる。下の図では、刺激電極をマイナスにした場合で、電流が赤い矢印のように流れ、細胞内部が過分極(hyperpolarization)になる。
電流の向きを逆にして、刺激電極で細胞膜を内側から外側に流れるような電流を短い時間与えると、細胞内が静止電位から0の方に変位する。これを脱分極(depolarization)と呼ぶ。 脱分極がおこると、これを平衡電位へ戻すようにカリウムイオンが細胞外へ移動する。一方、脱分極によって電位依存性のナトリウムチャンネルが開き、さらに脱分極を大きくする方向にナトリウムイオンが流入する。
脱分極の大きさがあまり大きくならないうちに刺激を止めると、平衡電位へ戻ってしまうが、脱分極の大きさが十分に大きくなり、閾値電位(threshold potential)を越えると、電位依存型のナトリウムチャンネル分子で「開」状態のものが多くなり、ナトリウムイオンがさらに流入し電位変化が起こり、これがさらにナトリウムチャンネルを「開」にする、というサイクルが動き出し、刺激を止めてももう元には戻らなくな、急激にNa+ の平衡電位に近づく。つまり電位は0ではなくプラスになる(オーバーシュートという)。
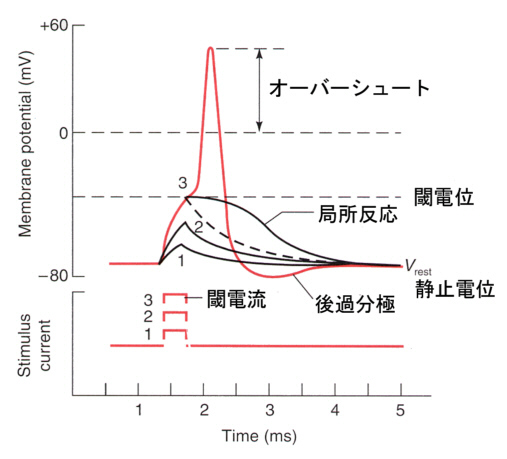
その後、電位依存性ナトリウムチャンネルが閉じると、少し遅れて電位依存性のカリウムチャンネル(静止電位の項で述べたカリウムチャンネルとは異なる)が開き、電位は再びK+の平衡電位に戻る。この一過的な電位の変化を活動電位と言う。
以上を動画にしたものが、次のページにある。
http://www.blackwellscience.com/matthews/channel.html
活動電位は、インパルス(impulse)とかスパイク(spike)と呼ばれることがある。
これまで述べたことは、人為的に電気刺激を加えたときの神経細胞の反応である。生体では、シナプスを通じて樹状突起や細胞体で受けた、さまざまな情報が統合され(この点に関しては後述)、軸索丘で活動電位が生じる。この部分には電位依存型のナトリウムチャンネルの数が多い。
さらに学びたい人へ
http://human.physiol.arizona.edu/SCHED/CV/Wright/15action.htm
http://www.unl.edu/tcweb/pharm/con.action.potential.html
軸索丘で発生したインパルスは、軸索を伝わってすばやく軸索末端まで伝えられ(伝導、conduction)、軸索の末端から放出される神経伝達物質(neurotransmitter)の放出という形で、信号がシナプスを形成している次の神経細胞あるいは筋肉に伝達される(transmission)。
活動電位が軸索を伝わり、しかも伝導の方向が常に細胞体側から軸索末端に向かう方向のみで、逆転が起こらないのはなぜなのだろうか。
HodgkinとHuxleyらの電気生理学的研究によって神経細胞の興奮とその伝導は、細胞膜のイオン透過性の変化に基づくことが明らかになり、細胞膜にあって膜電位の変化によって特定のイオンを選択的に透過させるイオンチャンネルの存在が示された。
軸索丘でのイオンチャンネルの開口によってナトリウムイオンが流入すると、その場所はナトリウムイオンの平衡電位に近づく(すなわち内側が正になる)。この結果、隣接する細胞膜(ここはまだ内側が負のままである)との間に局所回路が生じ、この局所回路が隣接部位を脱分極に導く。すると隣接部位のナトリウムチャンネルが開口し、その部位を正に導く。それが局所回路を形成し、また次の部位を興奮(excitation)させる。こうして局所回路とその結果生じるナトリウムチャンネルの開口の連鎖が次々と隣に伝わってゆく。
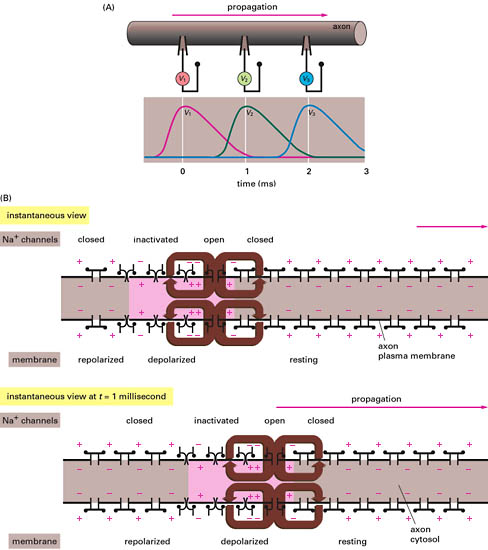
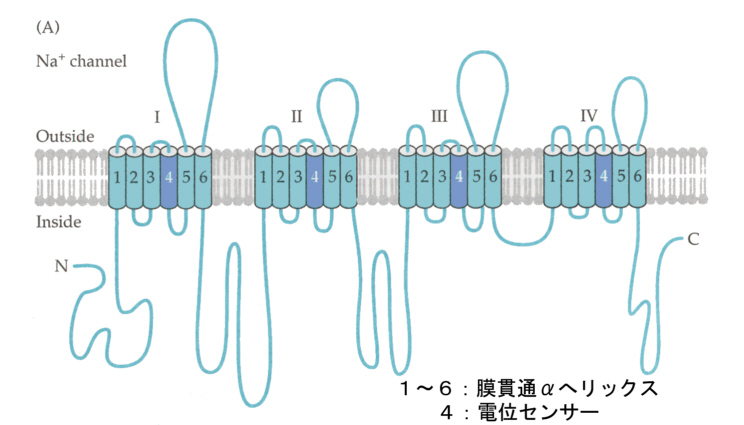
電位依存型のナトリウムチャンネルの構造は次のように考えられている(アミノ酸配列から推測したモデル)。大きな分子はローマ数字で表した4つの領域に分かれ、それぞれは細胞膜を6回貫通している。5と6のドメインから細胞外へ伸びたランダムコイル部分は親水性が高く、この部分が下側へ折れ込んで、チャンネルを形成している。ドメイン4は電荷を持ったアミノ酸(リジンとアルギニン)を含んでいてこれが電位センサーとして働いていると考えられている。膜の電位の変化に応じてこの電位センサー部位が動き、その結果4つの領域で作っているチャンネル(穴)が広がる。
膜の興奮が一方向のみに伝わるのは、一度活性化されたナトリウムチャンネルはすぐに不活性化され、その後すぐには活性化されない不応期があるからである。そのため、興奮は後戻りはできないのである。これはナトリウムチャンネルに、閉(次の図のBeginning)、開(次の図のDepolarization)、不活性化(次の図のInactivation)の3つの状態があるためだと考えられている。
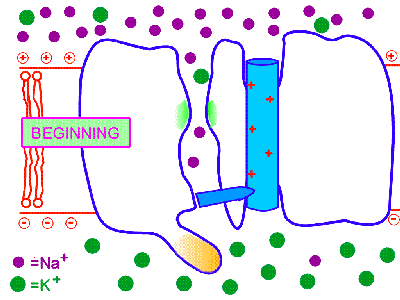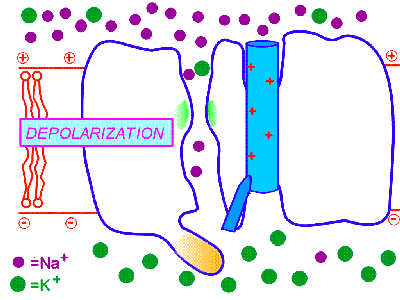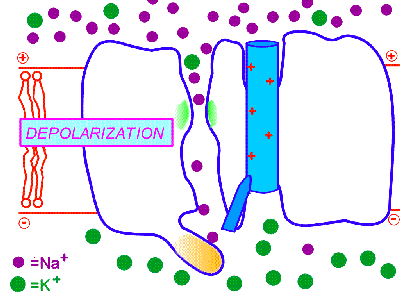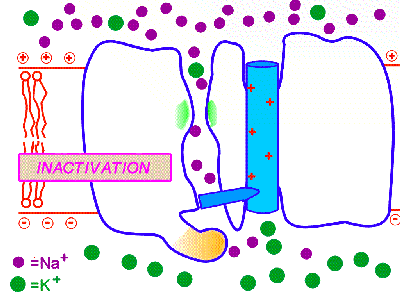
http://courses.washington.edu/conj/membrane/nachan.htmより
軸索丘から発射される活動電位の大きさは、刺激の大きさによらず常に一定である。刺激が大きくなると変化するのは単位時間あたりに発射されるインパルスの数である。したがって、刺激の強さというアナログ量はインパルスの頻度というデジタル量に変えられて伝導してゆく。
跳躍伝導 運動神経は有髄の軸索を持つ。解剖の時に見た座骨神経などの脊髄神経の束を思い出してほしい。白く光っていたはずである。これは髄鞘と呼ばれる脂質に富んだ、したがって絶縁性の高い部分が見えていたのである。髄鞘(ミエリン鞘)は軸索が伸びてゆくときに、シュワン細胞(中枢では稀突起グリア細胞の突起)がちょうど絶縁テープを巻くように軸索の回りに巻きついたものである。
髄鞘はずっと連続しているわけではなく、一定間隔(約1mm)ごとにとぎれるところがある。ここをランビエの絞輪とよぶ。ナトリウムイオンチャンネルはこの部分に分布しているので、前節で述べた局所回路は絶縁性の高い髄鞘を飛ばして次の絞輪との間に形成される。したがって伝導は飛び飛びにゆくので跳躍伝導と呼ばれる。伝導速度は無髄神経に比べてずっと早くなる。
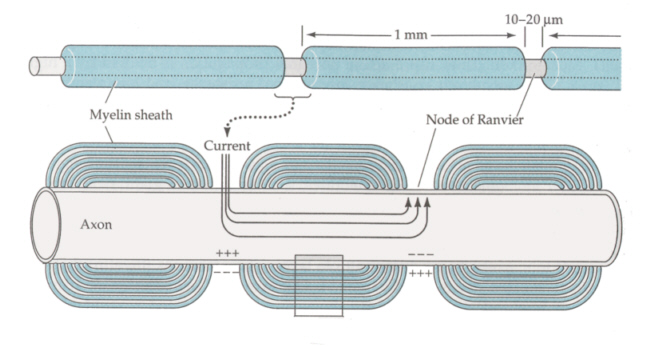
無髄神経と有髄神経の伝導を動画にしたものが、次のページにある。
http://www.blackwellscience.com/matthews/actionp.html
![]() この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
 ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。
ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。