
神経幹細胞の分化メカニズムの研究 (~2010)
わたしたちは神経幹細胞についても研究を行っています。わたしたちは20年ちかく前に未分化細胞特異的転写因子Oct-3(Oct-3/4, Oct-4と同じもの)を発見しました(東京大学第一生化学教室<村松正実教授>における濱田博司助教授グループの業績)。岡澤が学位研究として初めて行ったのがOct-3/4のクローニングであり、自らの手でcDNAと染色体クローンの両者を発見しました(Cell 1990; EMBO J 1991)。
研究の発想は、未分化状態のみに発現する転写因子が分化のスイッチになるだろうという予測でしたが、これは多くの研究者が分化後すぐに発現する転写因子を探していたのと逆の発想でした。スイッチ分子かどうかは、その後Oct-3/4のstable cell lineを用いた研究で、Oct-3/4発現と形態変化の相関を確認しましたが、完全な証明には至りませんでした(EMBO J 1993)。しかし、10年ほど経ってから、英国のSmith教授あるいはその共同研究者のNiwa博士らによって、私たちが妄想していたとおりにOct-3/4が胚性幹細胞(ES細胞)の分化のマスタースイッチとして機能することが分かってきました(Cell 1998, Nat Genet 2000)。さらに、そのような知見と関連して、Yamanaka教授らの研究から、Oct-3/4を4転写因子でiPS細胞が創出されました。その後、4因子の中でも、Oct-3/4が最重要であることが多くの論文から示唆されています。例えば、Oct-3/4単独でもiPS細胞が神経幹細胞から作成出来ることがマックスプランク研究所長のHans Schoeler博士らの研究から示されています(Nature 2010)。これらの、国内外で行われたすばらしい研究によってES細胞あるいはiPS細胞におけるOct-3/4の機能が証明されてきました。現在、わたくしたちは神経変性疾患の研究をする立場から神経幹細胞でのOct-3の機能を調べています。このことはやがて神経変性疾患の治療にも役立つ知識を与えてくれるものと考えています。
特に、最近私たちは神経幹細胞においてもOct-3/4が発現していることを発見しました(Okuda et al., Mol Brain Res 2004)。神経幹細胞においても、ES細胞同様にある種の分化のゲートキーパーとして働きうるのかということに興味を持っています。一方、神経幹細胞特異的にOct-3/4を欠失するconditional KOマウスを作製しても、なんら症状が起きない、との報告が最近なされ(Lengner et al., Cell Stem Cell 2007)、Oct-3/4が神経幹細胞に存在するのか?あるいは、以前に報告されたものはpseudogeneではないのか?という論争が起きています。そこで、私たちは精製した神経幹細胞からRNAを抽出し、RT-PCR産物(ゲノムDNAからは増幅されないようにして得られたもの)を直接シーケンスしてみました(Chin et al., BBRC 2009)。その結果、神経幹細胞も明らかにOct-3/4を発現しているとの結論が得られました。したがって、神経幹細胞におけるOct-3/4の機能については更なる検討が必要と考えられます。
研究の発想は、未分化状態のみに発現する転写因子が分化のスイッチになるだろうという予測でしたが、これは多くの研究者が分化後すぐに発現する転写因子を探していたのと逆の発想でした。スイッチ分子かどうかは、その後Oct-3/4のstable cell lineを用いた研究で、Oct-3/4発現と形態変化の相関を確認しましたが、完全な証明には至りませんでした(EMBO J 1993)。しかし、10年ほど経ってから、英国のSmith教授あるいはその共同研究者のNiwa博士らによって、私たちが妄想していたとおりにOct-3/4が胚性幹細胞(ES細胞)の分化のマスタースイッチとして機能することが分かってきました(Cell 1998, Nat Genet 2000)。さらに、そのような知見と関連して、Yamanaka教授らの研究から、Oct-3/4を4転写因子でiPS細胞が創出されました。その後、4因子の中でも、Oct-3/4が最重要であることが多くの論文から示唆されています。例えば、Oct-3/4単独でもiPS細胞が神経幹細胞から作成出来ることがマックスプランク研究所長のHans Schoeler博士らの研究から示されています(Nature 2010)。これらの、国内外で行われたすばらしい研究によってES細胞あるいはiPS細胞におけるOct-3/4の機能が証明されてきました。現在、わたくしたちは神経変性疾患の研究をする立場から神経幹細胞でのOct-3の機能を調べています。このことはやがて神経変性疾患の治療にも役立つ知識を与えてくれるものと考えています。
特に、最近私たちは神経幹細胞においてもOct-3/4が発現していることを発見しました(Okuda et al., Mol Brain Res 2004)。神経幹細胞においても、ES細胞同様にある種の分化のゲートキーパーとして働きうるのかということに興味を持っています。一方、神経幹細胞特異的にOct-3/4を欠失するconditional KOマウスを作製しても、なんら症状が起きない、との報告が最近なされ(Lengner et al., Cell Stem Cell 2007)、Oct-3/4が神経幹細胞に存在するのか?あるいは、以前に報告されたものはpseudogeneではないのか?という論争が起きています。そこで、私たちは精製した神経幹細胞からRNAを抽出し、RT-PCR産物(ゲノムDNAからは増幅されないようにして得られたもの)を直接シーケンスしてみました(Chin et al., BBRC 2009)。その結果、神経幹細胞も明らかにOct-3/4を発現しているとの結論が得られました。したがって、神経幹細胞におけるOct-3/4の機能については更なる検討が必要と考えられます。
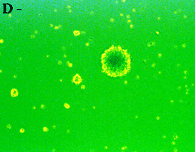

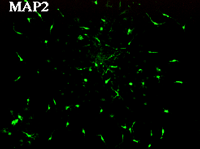
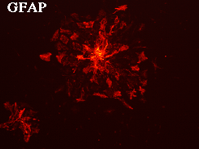
神経幹細胞(neurosphere,上段左)をpoly-L-lysineでcoatingした培養皿に移すと(上段右)、MAP2陽性の神経細胞(下段左)とGFAP陽性のグリア細胞(下段右)に分化する。上段右、下段左、下段右は同一視野。
発表論文
Okamoto K, Okazawa H, Okuda A, Sakai M, Muramatsu M, Hamada H.
A novel octamer binding transcription factor is differentially expressed in mouse embryonic cells.
Cell. 60(3) 461-472 1990. doi: 10.1016/0092-8674(90)90597-8.
Okazawa H, Okamoto K, Ishino F, Ishino-Kaneko T, Takeda S, Toyoda Y, Muramatsu M, Hamada H.
The oct3 gene, a gene for an embryonic transcription factor, is controlled by a retinoic acid repressible enhancer.
EMBO Journal 10(10) 2997-3005 1991. PMCID: PMC453014
Shimazaki T, Okazawa H, Fujii H, Ikeda M, Tamai K, McKay RD, Muramatsu M, Hamada H.
Hybrid cell extinction and re-expression of Oct-3 function correlates with differentiation potential.
EMBO Journal 12(12) 4489-4498 1993. PMCID: PMC413873
Okuda T, Tagawa K, Qi ML, Hoshio M, Ueda H, Kawano H, Kanazawa I, Muramatsu M, Okazawa H.
Oct-3/4 repression accelerates differentiation of neural progenitor cells in vitro and in vivo.
Molecular brain research 132 18-30 Dec 2004. doi: 10.1016/j.molbrainres.2004.08.021.
Chin JH, Shiwaku H, Goda O, Komuro A, Okazawa H.
Neural stem cells express Oct-3/4.
Biochemical and biophysical research communications 388(2) 247-251 Oct 2009. doi: 10.1016/j.bbrc.2009.07.165. Epub 2009 Aug 5.
A novel octamer binding transcription factor is differentially expressed in mouse embryonic cells.
Cell. 60(3) 461-472 1990. doi: 10.1016/0092-8674(90)90597-8.
Okazawa H, Okamoto K, Ishino F, Ishino-Kaneko T, Takeda S, Toyoda Y, Muramatsu M, Hamada H.
The oct3 gene, a gene for an embryonic transcription factor, is controlled by a retinoic acid repressible enhancer.
EMBO Journal 10(10) 2997-3005 1991. PMCID: PMC453014
Shimazaki T, Okazawa H, Fujii H, Ikeda M, Tamai K, McKay RD, Muramatsu M, Hamada H.
Hybrid cell extinction and re-expression of Oct-3 function correlates with differentiation potential.
EMBO Journal 12(12) 4489-4498 1993. PMCID: PMC413873
Okuda T, Tagawa K, Qi ML, Hoshio M, Ueda H, Kawano H, Kanazawa I, Muramatsu M, Okazawa H.
Oct-3/4 repression accelerates differentiation of neural progenitor cells in vitro and in vivo.
Molecular brain research 132 18-30 Dec 2004. doi: 10.1016/j.molbrainres.2004.08.021.
Chin JH, Shiwaku H, Goda O, Komuro A, Okazawa H.
Neural stem cells express Oct-3/4.
Biochemical and biophysical research communications 388(2) 247-251 Oct 2009. doi: 10.1016/j.bbrc.2009.07.165. Epub 2009 Aug 5.