
筋特異的選択的スプライシング
Nature Methods (2006), Mol Cell Biol (2007)
選択的スプライシングによってリガンド特異性の切替えが起こる線虫の線維芽細胞成長因子受容体遺伝子の筋特異的選択的スプライシングの制御機構について紹介します.
選択的スプライシングによってリガンド特異性の切替えが起こる線虫の線維芽細胞成長因子受容体遺伝子の筋特異的選択的スプライシングの制御機構について紹介します.
線虫線維芽細胞成長因子(FGF)受容体遺伝子
線虫のegl-15遺伝子は線虫で唯一のFGF受容体をコードする遺伝子で、相互排他的なエクソン5Aとエクソン5Bの使い分けによって、線虫に2種類存在するFGF(EGL-17およびLET-756)とのリガンド結合特異性が規定されます.
LET-756/EGL-15Bシグナルが線虫の生存に必須であるのに対し、EGL-17/EGL-15Aは性筋芽細胞の誘引因子とその受容体として機能し、egl-17変異体あるいはegl-15エクソン5A特異的変異体は、雌雄同体の産卵に必須の陰門筋が正常な位置で分化できず、産卵障害(egg-laying defective, Egl)の表現型を示します.
LET-756/EGL-15Bシグナルが線虫の生存に必須であるのに対し、EGL-17/EGL-15Aは性筋芽細胞の誘引因子とその受容体として機能し、egl-17変異体あるいはegl-15エクソン5A特異的変異体は、雌雄同体の産卵に必須の陰門筋が正常な位置で分化できず、産卵障害(egg-laying defective, Egl)の表現型を示します.
egl-15スプライシングレポーター
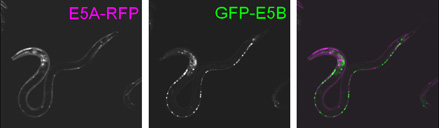
egl-15レポーター線虫(Nature Methods, 2006より)
蛍光タンパク質GFPとRFPを用いて、egl-15遺伝子のエクソン5Aとエクソン5Bの選択をモニターできるレポーターミニ遺伝子を作製しました.このレポーターでは、RFPの発現はエクソン5Aの選択を、GFPの発現はエクソン5Bの選択を反映します.
このレポーターを全身で発現させると、筋組織は5A型(RFP)を、上皮系や神経系などそれ以外の組織は5B型(GFP)を主に発現し、組織特異的選択性を示すことが分かりました(図).
このレポーターを全身で発現させると、筋組織は5A型(RFP)を、上皮系や神経系などそれ以外の組織は5B型(GFP)を主に発現し、組織特異的選択性を示すことが分かりました(図).
変異体の単離と制御因子の同定
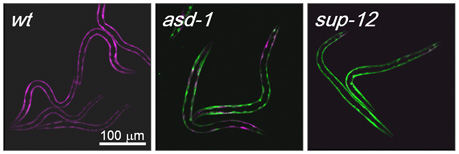
egl-15レポーターの変異体(Nature Methods, 2006より)
筋組織特異性の制御機構を解明するために、レポーターの色が異常になる変異体のスクリーニングを行い、筋における選択性がエクソン5A型から5B型に変化した変異体が多数得られました.
変異体の原因遺伝子のマッピングとクローニングの結果、2つのファミリーに属する3種類のRNA結合タンパク質ASD-1 (Alternative-Splicing-Defective-1,私たちの研究室が命名)、FOX-1およびSUP-12がegl-15遺伝子の筋組織特異的選択的スプライシングを制御していることを見出しました.
これらの制御因子の発現を解析すると、筋組織でのみ3者が共通して発現しており、腸や神経にはASD-1/FOX-1発現があるにもかかわらずegl-15のスプライシング制御には不十分であることが分かりました.
さらに、これらのスプライシング変異体は、egl-15(5A)特異的変異体やegl-17変異体と同じ性筋芽細胞の移動異常による陰門筋の形成不全と産卵障害の表現型を示したことから、これらのRNA結合タンパク質による組織特異的選択的スプライシング制御によって内在性FGF受容体のリガンド特異性が規定されていること、EGL-15のリガンド特異性の規定が、Fox-1ファミリーとSUP-12による組織特異的選択的スプライシング制御の重要な標的であることが遺伝学的に明らかとなりました.
変異体の原因遺伝子のマッピングとクローニングの結果、2つのファミリーに属する3種類のRNA結合タンパク質ASD-1 (Alternative-Splicing-Defective-1,私たちの研究室が命名)、FOX-1およびSUP-12がegl-15遺伝子の筋組織特異的選択的スプライシングを制御していることを見出しました.
これらの制御因子の発現を解析すると、筋組織でのみ3者が共通して発現しており、腸や神経にはASD-1/FOX-1発現があるにもかかわらずegl-15のスプライシング制御には不十分であることが分かりました.
さらに、これらのスプライシング変異体は、egl-15(5A)特異的変異体やegl-17変異体と同じ性筋芽細胞の移動異常による陰門筋の形成不全と産卵障害の表現型を示したことから、これらのRNA結合タンパク質による組織特異的選択的スプライシング制御によって内在性FGF受容体のリガンド特異性が規定されていること、EGL-15のリガンド特異性の規定が、Fox-1ファミリーとSUP-12による組織特異的選択的スプライシング制御の重要な標的であることが遺伝学的に明らかとなりました.
シスエレメントの同定

3種の線虫のegl-15遺伝子イントロン4の配列
筋特異性の制御に必要なシスエレメントの同定も、レポーターミニ遺伝子を用いて行いました.
全ゲノムの塩基配列が明らかにされている3種の線虫C. elegans、C. briggsae、C. remaneiで、相互排他的なエクソン5Aと5Bの構造が保存されています.そして、筋特異的に抑制される非筋型エクソン5Bの上流のイントロン4に、線虫類に保存された2つの配列UGCAUGとGUGUGが保存されています.ミニ遺伝子のこれらの配列にそれぞれ変異を導入しトランスジェニック線虫を作製したところ、スプライシング変異体同様に筋におけるエクソン5A選択性に異常を示して5A型から5B型に変化したことから、これらの配列が筋組織特異性に必要なシスエレメントと同定されました.
さらに、ゲルシフト実験および紫外線架橋による試験管内結合試験で、ASD-1/FOX-1とSUP-12はUGCAUGとGUGUGにそれぞれ特異的に結合すること、両タンパク質が同時に存在することによって協調的に標的RNAに結合することを確認しました.
全ゲノムの塩基配列が明らかにされている3種の線虫C. elegans、C. briggsae、C. remaneiで、相互排他的なエクソン5Aと5Bの構造が保存されています.そして、筋特異的に抑制される非筋型エクソン5Bの上流のイントロン4に、線虫類に保存された2つの配列UGCAUGとGUGUGが保存されています.ミニ遺伝子のこれらの配列にそれぞれ変異を導入しトランスジェニック線虫を作製したところ、スプライシング変異体同様に筋におけるエクソン5A選択性に異常を示して5A型から5B型に変化したことから、これらの配列が筋組織特異性に必要なシスエレメントと同定されました.
さらに、ゲルシフト実験および紫外線架橋による試験管内結合試験で、ASD-1/FOX-1とSUP-12はUGCAUGとGUGUGにそれぞれ特異的に結合すること、両タンパク質が同時に存在することによって協調的に標的RNAに結合することを確認しました.
制御のモデルと研究成果の意義
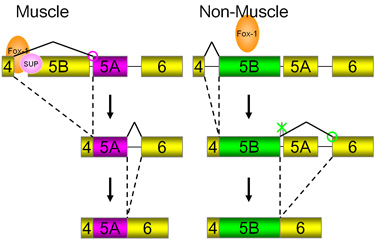
egl-15遺伝子の筋特異的制御のモデル
筋では、ASD-1/FOX-1とSUP-12が協調してイントロン4に結合してエクソン5Bを抑制し、下流のエクソン5Aが選択されます(図左).制御因子の変異体ではこの抑制が不十分で、エクソン5Bが選択されます(図右).
この研究結果は、複数のシスエレメントとそれらに結合する複数の制御因子がすべてそろうことで初めて効率的にエクソンの選択性が制御されることを示しています.制御因子の標的エクソンとその選択の組織特異性が複数のシスエレメントと複数の制御因子の組み合わせにより厳密にされていることを示す好例となりました.
また、スプライシングレポーターの色が内在性の遺伝子のスプライシングパターンを反映し、レポーターの色の異常を指標に変異体スクリーニングすることで内在性の遺伝子の選択的スプライシングの新規の制御因子を同定できたことから、私たちの生体内選択的スプライシングレポーターがスプライシング暗号解明のための有用なツールであることが示されました.
脊椎動物においてもFox-1ファミリー(RBFOX1, RBFOX2, RBFOX3)がUGCAUG配列を介して組織特異的選択的スプライシングを制御していることが多数報告されています.私たちの線虫egl-15遺伝子制御機構の研究成果により、Fox-1ファミリーとUGCAUGによる組織特異的選択的スプライシング制御機構が脊椎動物から線虫まで進化的によく保存されていることが明らかとなりました.
哺乳類のFGF受容体FGFR2遺伝子の組織特異的選択的スプライシングもFox-1ファミリーによって制御されています.Fox-1ファミリーがUGCAUGを介してFGF受容体の相互排他的選択的スプライシングを制御してリガンド特異性を規定するという構図が哺乳類と線虫で相似形をなしており、進化的な面からもたいへん興味深い事例です.
この研究結果は、複数のシスエレメントとそれらに結合する複数の制御因子がすべてそろうことで初めて効率的にエクソンの選択性が制御されることを示しています.制御因子の標的エクソンとその選択の組織特異性が複数のシスエレメントと複数の制御因子の組み合わせにより厳密にされていることを示す好例となりました.
また、スプライシングレポーターの色が内在性の遺伝子のスプライシングパターンを反映し、レポーターの色の異常を指標に変異体スクリーニングすることで内在性の遺伝子の選択的スプライシングの新規の制御因子を同定できたことから、私たちの生体内選択的スプライシングレポーターがスプライシング暗号解明のための有用なツールであることが示されました.
脊椎動物においてもFox-1ファミリー(RBFOX1, RBFOX2, RBFOX3)がUGCAUG配列を介して組織特異的選択的スプライシングを制御していることが多数報告されています.私たちの線虫egl-15遺伝子制御機構の研究成果により、Fox-1ファミリーとUGCAUGによる組織特異的選択的スプライシング制御機構が脊椎動物から線虫まで進化的によく保存されていることが明らかとなりました.
哺乳類のFGF受容体FGFR2遺伝子の組織特異的選択的スプライシングもFox-1ファミリーによって制御されています.Fox-1ファミリーがUGCAUGを介してFGF受容体の相互排他的選択的スプライシングを制御してリガンド特異性を規定するという構図が哺乳類と線虫で相似形をなしており、進化的な面からもたいへん興味深い事例です.