生物ⅠB(第一学習社)より
0.はじめに 1.動物の生殖 2.配偶子形成 3.受精 4.初期発生 5.器官形成 6.始原生殖細胞と性分化 関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください) University of Virginia, Human Biology Sect3 Development http://www.med.unc.edu/embryo_images/unit-welcome/welcome_htms/contents.htm http://www.ucalgary.ca/UofC/eduweb/virtualembryo/db_tutorial.html http://www.uoguelph.ca/zoology/devobio/dbindex.htm 更新日:2001年11月16日
最後の2回で動物の発生について学ぼう。
発生学(embryology, developmental biology)は、生物学の中でも昔からある学問分野である。ヒトが生物として、個体の誕生や成長、その他の動物、たとえばカエルの卵がオタマジャクシになって、成体になるような現象を見て、不思議に思ってきたからである。どうして均一に見えるカエルの卵から、複雑な形態をしたオタマジャクシが泳ぎだし、カエルになるのだろうか。この誰もが不思議に思う現象を、解き明かそうとするのが発生学である。
発生学は、上にカッコ書きをした英語からわかるように、始めはembryo、すなわち胚(ヒトでは胎児)がどう変化していくかを記述するところから始まった。それが発生(development)に伴うさまざまな現象を生物学のあらゆる手法で、どうしてそうなるのかを理解していく方向が鮮明になり、英語の名前もそれを反映するようなものに変わった。
したがって、現在の発生学を理解するためには、これまで学習したあらゆる知識が必要である。たとえば、
遺伝子とその発現機構
タンパク質への翻訳機構
細胞と細胞の接着、組織の構築
いろいろな器官系の構造
体細胞分裂と減数分裂
細胞周期とその調節
転写調節機構
細胞間の情報交換の機構(リガンド、受容体、細胞内のシグナル伝達機構)
など、などである。もう一度見直して知識を確実にしておいて欲しい。
ここでは、少し広い範囲から話を始めよう。
個体の一生は限られたものなので、どこかで必ず次の世代を作って種の存続をはかっている。すなわち生殖(reproduction)である。生殖は、個体維持とならんで最も生物らしい活動である。
生物界全体を見渡すと、無性生殖(asexual reproduction)と有性生殖(sexual reproduction)が区別できるが、動物界には無性生殖をおこなう種はそれほど多くはない。
動物の無性生殖の例としては、次のようなものが挙げられる。
出芽(budding):ヒドラ
細片分離あるいは断烈(fragmentation):腔腸動物、扁形動物
単為生殖(parthenogenesis):ミツバチ、アリマキ
無性生殖というのは、自分と全く同じ遺伝子を持った個体を新たに作り出すわけで、安定した環境ではどんどん増えていけばいいので好都合だが、環境が変化したときには対応することはできない。
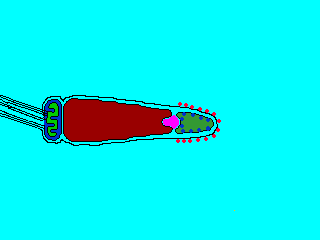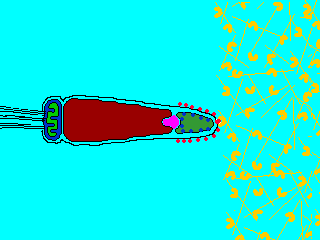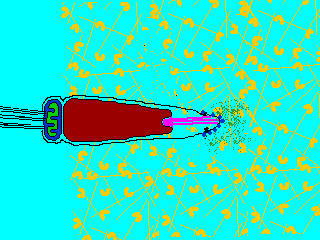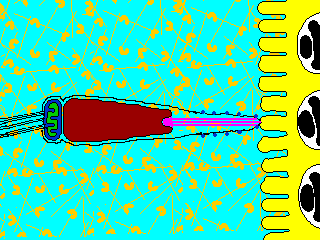
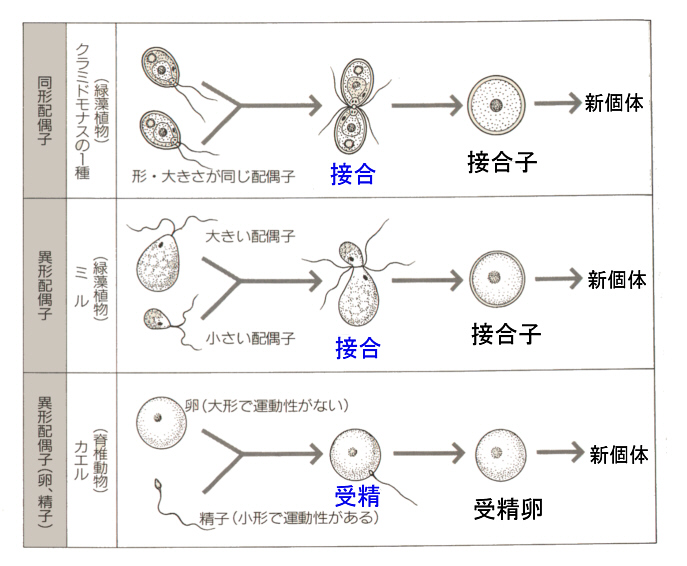
そこで進化の過程で作り出されたのが、遺伝子を混ぜ合わせる有性生殖(sexual reproduction)である。有性生殖では、雄と雌で作られた生殖細胞、すなわち配偶子(gamate)が融合して、新しい個体が生じる。配偶子には形・大きさが同じ同型配偶子をつくる生物と、配偶子の大きさに差がある異型配偶子をつくる生物がいる。雌雄の作る異型配偶子に、形態・分化に大きな差が見られるとき、雌のものを卵(ovum)、雄のものを精子(spermatozoon)という。ふつうは、卵は大きく運動性を欠き、精子は小さく運動性がある。動物は卵と精子を作る場合がほとんででる。
生物ⅠB(第一学習社)より
精子と卵の癒合を受精(fertilization)といい、受精が雌の体内で起こる場合を体内受精(internal
fertilization)、体外で起こる場合を体外受精(external
fertilization)という。ちなみに配偶子どうしの融合は、接合と呼んでいる。
融合するのだから、染色体の数は配偶子の倍になる。そこで、配偶子を作るためには染色体の数を半減させておかなければならない。そのために、体細胞分裂とは異なる、減数分裂という、染色体を半減させる特別な分裂様式が導入された。
こうして大型の運動性のない配偶子である卵を作る雌と、運動性のある小型の配偶子である精子を作る雄の区別ができた。性(sex)とは、一般的には同種の生物に雄と雌の区別があることを言うが、もっとも根源的な意味は、雌雄の染色体の新しい組み合わせを作って、遺伝子を混ぜることにより、新しい組み合わせの遺伝子をもった個体を作ることである。日常会話では、この混ぜ合わせに必要な多くの現象(形態や行動を含めて)を性と呼ぶことが多い。
有性生殖を行うための器官を生殖器官と言い、配偶子を形成する生殖巣(生殖腺)とそれを体外へ排出するための生殖輸管、さらに生殖輸管に付属する腺などに分けられる。
雄の生殖腺は精巣(testis、pl. testes)であり、生殖輸管は輸精管(vas deferens)と呼ぶ。雌の生殖腺は卵巣(ovary)、生殖輸管は輸卵管(oviduct)、子宮(uterus)である。
生殖輸管の体外への出口を生殖口というが、体内受精を行う動物ではこの部位が交尾器(雄の陰茎など)として発達している。
1)減数分裂
減数分裂の過程は、体細胞分裂と比較しながらすでに1年生で学んだ(減数分裂の過程参照のこと)。
減数分裂の過程は、減数分裂Ⅰと減数分裂Ⅱの過程に分けられる。
Ⅰのとき相同染色体が対合する。この時、染色体はすでに複製されていて、対合の結果、父方の2本と母方の2本の合計4本の染色分体が並列し、染色分体の交叉と組み替えがおこる。その結果、染色分体は混ぜ合わされる。Ⅰの終期には染色分体は2本づつ分離し、染色体の数は半減する。
Ⅱになると、2個に分かれたそれぞれの細胞の染色分体が縦裂し、体細胞分裂と同じような過程でそれぞれの細胞に染色分体を分配する。こうして染色体数が半減した4つの生殖細胞が生じる。
ただし、実際の生殖細胞である精子と卵ができあがる過程は、もうすこし複雑である。精子と卵ができる過程は、それぞれ精子形成と卵形成と呼んでいる。以下に、脊椎動物のそれぞれの過程を述べる。
2)精子形成(spermatogenesis)
http://www.uoguelph.ca/zoology/devobio/210labs/testes1.html
成熟した精巣の断面を顕微鏡で観察すると、たくさんの丸い管の断面が見える。これは精細管(seminiferous tubule)で、この管は精巣の中をうねりながら最後は精巣網につながり、さらに精巣上体(epididymis)へとつながる。
精子は精巣上体から輸精管を経て生殖口へ出される。精細管どうしのあいだは結合組織で満たされ、毛細血管の断面が見え、間細胞(ライディッヒ細胞)と呼ばれる細胞も見られる。
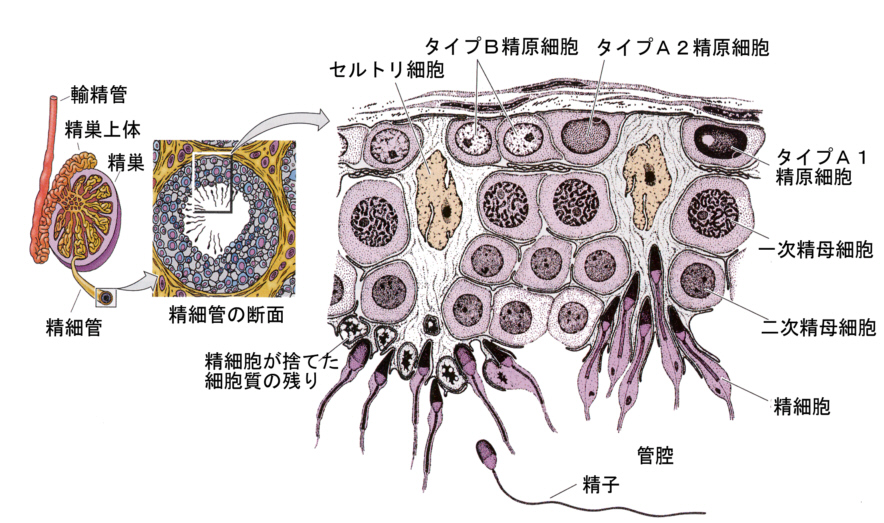
Developmental Biology, 6th ed. Scott Gilbert Sinauer Associates, Incより
精細管の断面はいろいろな大きさの細胞が詰まっていて、管腔に近い方には精子が存在する。精細管には二種類の細胞がある。1つは生殖細胞で、精子になるべき細胞であり、もう1つはセルトリ細胞で生殖細胞に栄養を補給し、また精子の停泊場となる。
精細管の一番内側からは、管腔に向かってセルトリ細胞が立ち上がるように配列している。基底膜に近いところには精子形成の出発点となる精原細胞(spermatogonium)が並んでいる。精原細胞は絶えず分裂して、精原細胞のストックを作るとともに、次ぎの段階へ移行する精原細胞となって行く。この過程は血球の発生のところで学習したのと同じ、幹細胞による増殖である。
精原細胞はさらに分裂して一次精母細胞(primary spermatocyte)になる。ここまでは体細胞分裂の過程である。はじめ核も細胞質も小さいが、時間をかけて成熟し、核が大きくなる。
ここから先が減数分裂の過程である。一次精母細胞は減数分裂Ⅰによって2個の二次精母細胞になり、続いておこる減数分裂Ⅱで4個の精細胞(spermatid)になる。
ここから先は精子に変態する過程である。精細胞は核も細胞質も小さく、セルトリ細胞の細胞質内に頭を突っ込んでいる。やがて精細胞は変態を始め、細胞質を失い、先体(acrosome)とそれに続く核を含む頭部、ミトコンドリアの詰まった中節、尾を備えた精子(sperm)となる。この過程で、精子が必要とする機能、すなわち卵へ侵入するための装置、遺伝情報、運動性だけを残し、あとは捨て去ってしまう。
電子顕微鏡による観察によると、次の段階へ移行する運命にある精原細胞は、分裂しても細胞質が完全には分離せず、細胞間橋によりつながっており、このつながりは以後、精細胞になるまで保たれる。そのため多数の細胞が同調的に分裂して一つながりの精子を作ることになる。
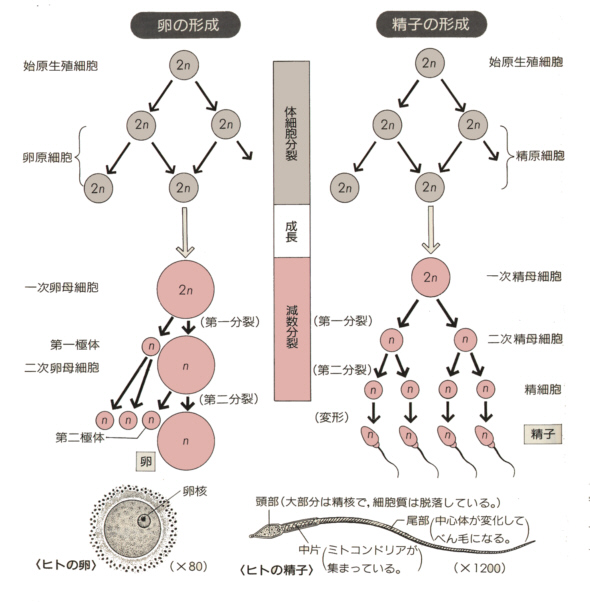
生物ⅠB(第一学習社)より
3)卵形成(oogenesis)
http://www.uoguelph.ca/zoology/devobio/210labs/ovary4.html
生殖器官は普通、左右一対あるが、両生類と鳥類の卵巣は左側のみが発達し、右側は痕跡的である。輸卵管も左側のみが発達する。成熟した卵巣は表面から見てもはっきり分かる多量の卵黄を蓄積した卵胞がブドウの房のように付いている。
卵も精子と同じような過程を経て作られるが、ヒトの場合、胎生期にさかんに生殖細胞は細胞分裂をして、多数の卵原細胞(oogonia)をつくられる。卵原細胞は体細胞分裂によって一次卵母細胞となる。ここからが減数分裂の過程であるが、卵の場合は細胞質を均等に分けるようなことはせず、著しく細胞質の少ない極体を放出して、第二次卵母細胞になる。第二次卵母細胞は減数分裂Ⅱの過程を経て卵となる。この時、極体も小さいながら分裂する。したがって最終的に、1つの卵と3つの極体が作られることになる。
ただし多くの動物で、減数分裂の過程は上に述べたように完全には終わらず、途中で止まっている。哺乳類では減数分裂Ⅱの中期で止まっている。
精子が侵入すると、止まっていた減数分裂が再開し、極体を放出して染色体数の半減した卵となる。
出生後は卵原細胞の分裂は停止し、これを一層の卵胞上皮細胞が包む。これを一次卵胞(原始卵胞)とよぶ。ヒトの新生児や幼児では卵胞の総数は50万個と言われる。思春期には約16万個になる。その後、月経周期あたり1個の卵胞のみが成熟して排卵する。一生のうちには結局400個足らずの卵が排卵されるのみで、他の大部分は十分成熟せず排卵もされず退化する。
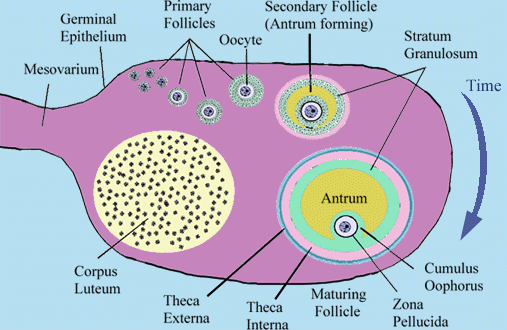
http://www.uoguelph.ca/zoology/devobio/210labs/ovary4.htmlより
成熟した雌の卵巣内には、多数の発達の程度の異なる卵胞(ovarian follicles)が存在する。一番小さい原始卵胞は生殖腺刺激ホルモン(LH、FSH)の作用によって卵胞上皮細胞が増殖し、やがて、卵胞液を貯留し、グラーフ卵胞となる。
成熟したグラーフ卵胞は、排卵前に起こるLHの一過的な大量分泌(LHサージ)によって、卵細胞を排卵する(ovulation)。哺乳類では、繁殖期にこの排卵が決まった間隔で、周期的に起こる。
卵の場合は、精子とは異なり、発生の過程に必要な栄養分とエネルギー源を蓄えておく必要がある。卵黄には卵黄タンパク質と中性脂肪が多量に蓄えられているが、これらはエストロジェンの作用によって肝臓で作られ、血液中を運ばれて卵巣に達するのである。哺乳類では胎生の発達により卵黄量は少ないが、鳥類や爬虫類では卵黄量の多い卵を作る。
卵にはこの他、多くの分子があらかじめ用意されている。
●リボソームとtRNA
●mRNA
●分化促進因子群
受精(fertilization)によって個体の一生が始まる。精子から持ち込まれた父方の遺伝情報と卵にある母方の遺伝情報が一緒になり、これまで地球上に存在しなかった新しい組み合わせが生まれる。一方、受精は最初の細胞間の認識でもある。
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
ウニの発生(模式的だが動画を含みわかりやすい)
受精が引き金になって、発生(development)が始まる。受精卵は卵割(cleavage)を始め、次第に細胞の数を増やしてゆく。受精によって発生の過程が始まるとともに、精子は中心体を卵内に持ち込み、分裂装置を作れるようにする。
卵割の様式やその後の発生の様子は動物によって異なる。この点に関しては、筋肉の発生の前置きとして第2回の講義で説明した(第2回の講義の内容スライド2から11参照のこと)ので、くわしいことはここでは触れない。
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
Developmental
Biology on Line (University of Guelph)
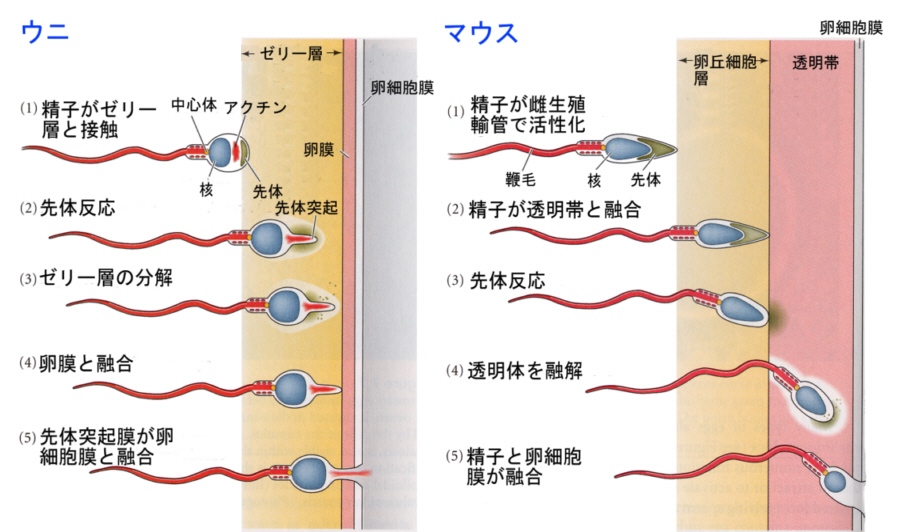
1)先体反応
精子は卵に誘引され、卵に近づき受精する。受精の過程を細かく見ると、先体反応が起こり、先体膜に埋め込まれたタンパク質(ウニの場合はバインディン)と卵細胞膜にある受容体との結合である。こうして同種の精子と卵が認識しあって、細胞膜が融合し、精子の頭部に納められていた核(正しくは雄性前核)と中心体が卵のなかに送り込まれる。
Developmental Biology, 6th ed. Scott Gilbert Sinauer Associates,
Incより
ウニの場合は先体反応によって先体突起が飛び出すが、哺乳類の場合は先体反応によって酵素を放出し、これが透明帯を溶かして精子が卵細胞膜へ到達するのを助ける。
精子細胞膜が卵細胞膜と融合できるのは、精子の先端にある卵の受容体に結合できる膜タンパク質のためである。ウニの場合、このタンパク質はバインディンと呼ばれている(上の動画の青い点)。バインディングと受容体に種特異異性があるので、別の種の精子による交雑を防ぐことができる。
2)多精拒否機構
一つの卵には一つの精子だけが入り、遅れたものは入ってもらっては困る。精子が複数卵に入る現象を多精と言うが、こうなると発生は正常には進まない。
そのため、卵には多精拒否機構(rejection of polyspermy)が備わっている。多精拒否機構には、早い機構と遅い機構の2つがある。
早い多精拒否機構は、精子が融合した後、1ないし3秒以内に完了する機構である。未受精卵にガラス電極を刺入して電位を測ると、内側が-70mVほどの電位が測定できる。これはニューロンのところで学習した静止電位に相当する。
精子が融合すると、すぐに+20mVほどに電位が上昇する。この電位は、活動電位と同じようにナトリウムイオンが関係している。つまりニューロンのように卵は精子と融合すると活動電位が発生するのである。この電位によって次に述べる受容体の立体構造が変化して、精子が結合できなくなると考えられている。
遅い多精拒否機構は、受精膜が挙がることによる。ウニを観察していると、精子が侵入したところから徐々に、受精膜が挙がり、やがて卵全体を包むようになる。この受精膜は、卵細胞膜のすぐ下に分布していた表層顆粒が細胞膜と融合し、その内容物を卵膜と細胞膜の間に放出することによって作られる。表層顆粒の内容物は卵膜と卵細胞膜の間に水を引き込み、すきまを押し広げるとともに、裏打ち構造を卵膜に付加して、卵膜を固い受精膜に変えている。

3)卵の賦活(活性化)
父方の遺伝情報を卵に送り込むのが精子の重要な役目であるが、もう一つ重要な役目がある。それは精子による卵の賦活である。精子が卵と融合して初めて発生の過程が動き出す。それはどうしてなのだろうか。
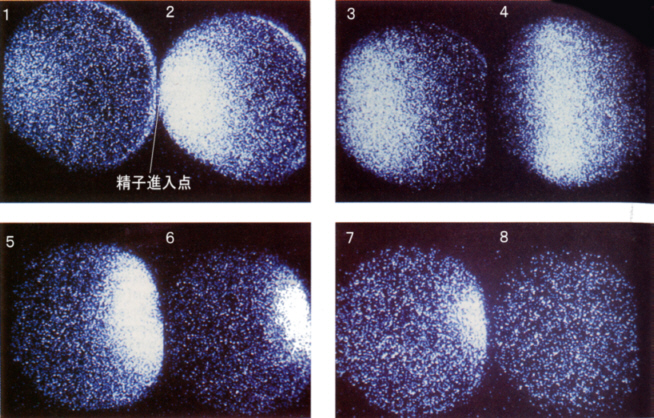
受精のとき、精子の侵入点から卵の表面を伝わって何かが伝播するという考えが以前からあって、これを受精波と呼んでいた。カルシウムが結合すると光るタンパク質が見つかり、これを卵内にあらかじめ注射をしておいて受精させると、精子の侵入点から光が卵全体に広がっていくことが観察された。受精波はカルシウム波だったのである。
Developmental Biology, 6th ed. Scott Gilbert Sinauer Associates,
Incより
精子が侵入してからカルシウム波が卵全体に広がっていくまでに、およそ30秒かかる。下の図の右側はカルシウム波、左側は受精膜が挙がるところを同時に示してある。
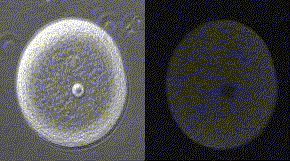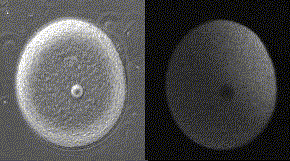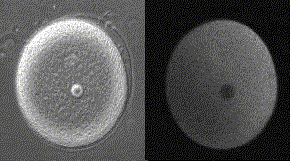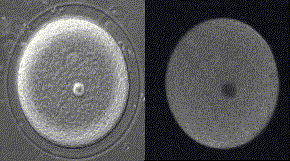
メダカ卵のカルシウム波(QuickTimeが必要)
http://sdb.bio.purdue.edu/dbcinema/jaffe/jaffe.html
つまり受精によって、卵内のカルシウムイオンが一過的に高まるのである。現在までの研究で、バインディンが受容体に結合すると、フォスフォリパーゼCが活性化され、イノシトール三リン酸が作られ、これがカルシウム貯蔵部位に作用してカルシウムイオンを放出させることがわかってきた。ホルモンと受容体の関係によく似ていることに驚くばかりである。
フォスフォリパーゼCが活性化にチロシンキナーゼが関与していることはわかっているが、受容体とチロシンキナーゼとフォスフォリパーゼCの間の関係はよく分かっていない。次のような機構モデルが考えられている。
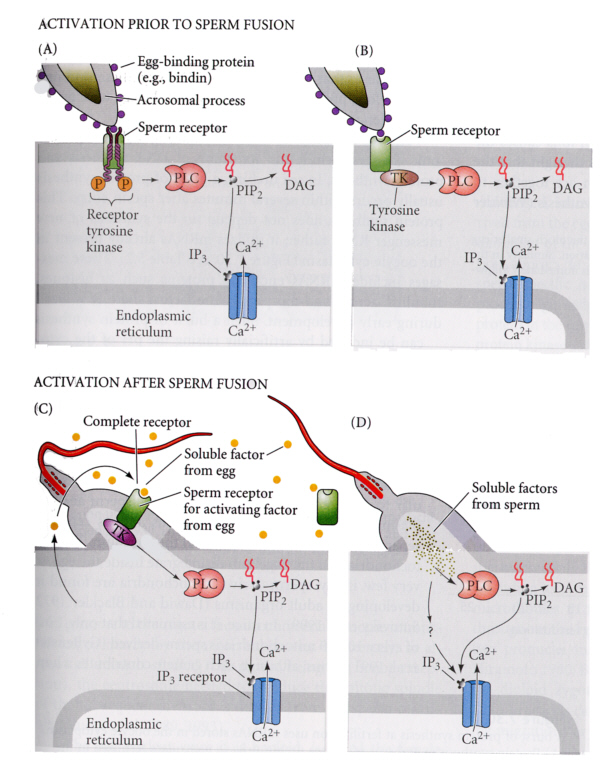
Developmental Biology, 6th ed. Scott Gilbert Sinauer Associates,
Incより
こうして、精子の持ち込んだ中心体から作られた分裂装置を使って、いよいよ卵割が始まるのである。
1)ウニの場合
初期発生は、卵割によって進んでいく。ウニを例に簡単に記すと、受精卵→2細胞期→4細胞期→8細胞期→16細胞期→桑実胚期→胞胚期→嚢胚期と進む。
胞胚から嚢胚になるときに陥入がおこり、一層だった細胞層が二重になり、細胞層の相互作用が可能となる。陥入を起こした部分を原口と呼び、貫入した部分を原腸という。原腸の先端は後に口を作り、原口は肛門となる。
陥入に前後して、胞胚内部に細胞群が落ち込む。これを一次間充織(primary mesenchyme)がいう。一次間充織は後の骨を作る。陥入が進み、原腸が胞胚腔の上端(貫入した場所の反対側)に達する頃に、原腸の先端から別の細胞群が胞胚腔に落ち込む。これを二次間充織(secondary mesenchyme)という。これらの間充織は中胚葉である。こうして外側の外胚葉(ectoderm)、原腸の内胚葉(endoderm)、間充織の中胚葉(mesoderm)という、3つの胚葉が分化する。

生物ⅠB(第一学習社)より
ウニでは発生が進むにつれて各胚葉から器官が形成され、胚の形が変化してプルテウス幼生になる。プルテウスはプランクトン生活を続けたあと、変態してウニの生体となって、着生生活を送るようになる。
ウニの発生についてさらに学びたい人へ
http://worms.zoology.wisc.edu/urchins/suwelcome.html
2)カエルの場合
脊椎動物のカエルの卵は端黄卵で、卵割様式は全割であるが、ウニの場合のように等割ではなく、動物極では割球が小さく、植物極では大きい不等割である。胞胚まではウニの場合とだいたい同じだが、胞胚後期になると、黒色の細胞が増えて、赤道面の下まで広がり、胞胚腔は卵黄の少ない動物局側にかたよった位置に生じる。
やがて植物極側(植物極点ではなく赤道面に寄ったところ)に半月状の切り込みができ、この溝を通って外側の細胞が内部へ侵入していく陥入が始まる。この溝が原口である。原口の動物極側(原口背唇)の細胞群は増殖しながら内部へ陥入を続けるとともに、動物極側へも移動し、動物極を覆い、さらに赤道面を越えて植物極側をも覆うようになる。
こうして外側を覆おう細胞群は外胚葉となり、内部に包み込まれた卵黄を含む細胞群は内胚葉となり、侵入した細胞群は中胚葉となる。
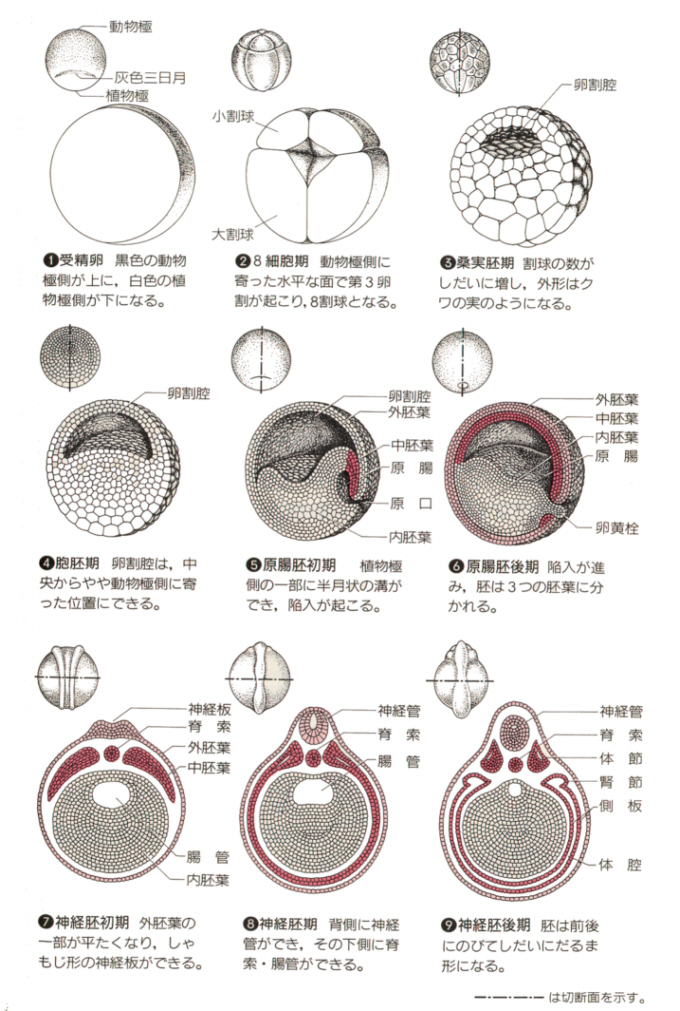
生物ⅠB(第一学習社)より
発生がさらに進むと、外胚葉は胚の表面を覆う表皮と背側を前後に走る神経管に分化する。中胚葉は、神経管の下側に沿って走る脊索と、脊索の両側の部分に分化し、この部分は背側から順に、体節、腎節、側板に分化する。
両生類の発生についてさらに学びたい人へ
http://worms.zoology.wisc.edu/frogs/mainmenu.html
ヒトの発生についても学びたい人へ
http://anatomy.med.unsw.edu.au/cbl/embryo/Embryo.htm#start
発生がさらに進むと、各胚葉からはいろいろの器官が分化する。カエルの場合は、外胚葉からは神経管が分化すると書いたが、神経管は後に脳と脊髄となる。この他の胚葉がどのような器官に分化するかを書いてみよう。
●外胚葉からできる器官
表皮 皮膚の表皮(毛、つめ、汗腺など)、眼の水晶体、角膜、口腔上皮、嗅上皮
神経管 脳、脊髄、脳神経、眼の網膜
●内胚葉からできる器官
消化管(食道・胃・小腸・大腸の内面の上皮)、えら、中耳、肺、気管
●中胚葉からできる器官
脊索(みずからは器官を作らないが、脊椎骨や筋肉の分化に関与する)
体節 脊椎骨、骨格、骨格筋(横紋筋)、皮膚の真皮
腎節 腎臓、輸尿管、生殖腺、生殖輸管(輸精管、輸卵管)
側板 腹膜、腸管膜、内蔵筋(平滑筋)、心臓、血管、結合組織
これは脊椎動物に共通している。
実際には、各胚葉はそれぞれが置かれた場所によって、いろいろな器官に分化していく。このような過程を器官形成(organogenesis)と言う。心臓の形成や腎臓の形成、あるいは四肢の形成など興味深い過程であるが、ここでは省略する。
カエルの初期器官形成をさらに学びたい人へ
http://www.uoguelph.ca/zoology/devobio/splab3/
どんな生物でも、発生が進み成熟すると雌では卵巣(ovary)、雄では精巣(testes)(両方をまとめて生殖腺 gonadと言う)が発達し、その中に卵と精子がそれぞれつくられ、次の受精へとつながってゆく。このように生殖細胞は、連綿として連なっていると考えることができる。生殖細胞を中心に考えれば、我々の身体は生殖細胞を過去から未来へ受け渡す運び屋に過ぎないとも考えられる。
身体を構成する器官は、すでに述べた三胚葉から分化する。じつは、三胚葉が分化する前に、これらの細胞群とは別に、将来、生殖細胞になる細胞群が分化する。そこでこれらの細胞の系譜を生殖細胞系列(germ cell line)と呼び、体細胞系列(somatic cell line)とは区別することがある。
器官形成で述べたさまざまな器官と異なり、生殖腺のできかたはやや特殊である。生殖腺は中胚葉からできる器官だと5.に述べているが、生殖細胞である精子や卵のもとになる細胞の起源は別なのである。
1)極細胞
ショウジョウバエのような昆虫の仲間の卵割は、表割という形式をとる。卵細胞質の分裂が起こらないで、核だけが分裂して数を増やしてゆく。産卵後およそ80分後、核が256個になった頃、核は表面に移動し始める。
さらに産卵後130分、核が512個になる頃には300ほどの核が 卵の表面に並び、後端には細胞質に包まれた細胞群があらわれてくる。この細胞群は極細胞(pole cell)と呼ばれ、後に生殖細胞となる。核の間に仕切りが生じて、一層の細胞層になるのはさらにずっと後の14回目の分裂の時である。
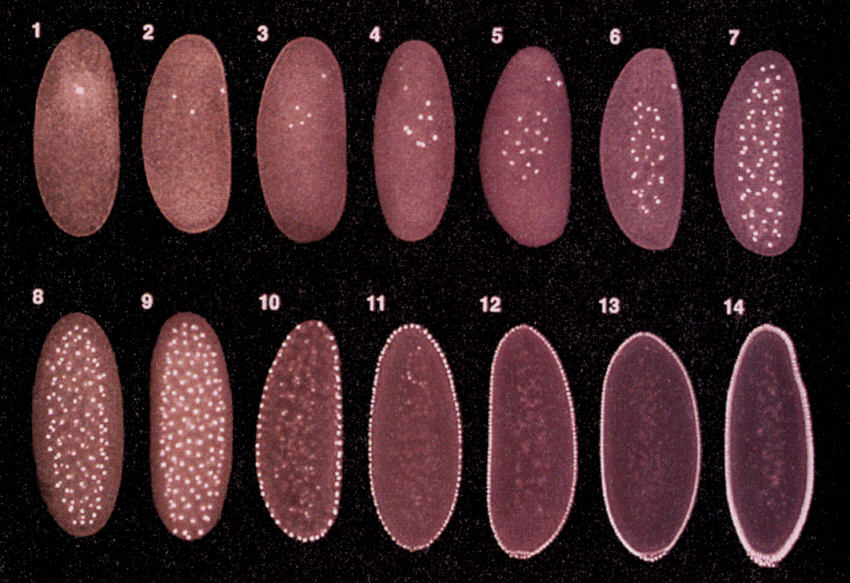
Developmental Biology, 6th ed. Scott Gilbert Sinauer Associates, Incより
このように将来、生殖細胞になる細胞は発生の比較的早い時期に、体細胞系列とは別の道をたどり始める。どうしてこのようなことが起こるのだろうか。
ショウジョウバエ卵の後端の細胞質を調べてみると、極顆粒と呼んでいる明瞭な顆粒が細胞質に存在する。後端の細胞質は他とは違っているのである(極細胞質)。この顆粒はミトコンドリア由来のRNAとタンパク質の複合体で、そこにやってきた核に働きかけて将来の運命を左右するらしいことはわかっている。
http://www.jtnet.ad.jp/WWW/JT/Culture/BRH/BRHTour/1F/book/97/16/library/scientist-1.html
卵細胞におけるこのような不均質性(あるいは極性とも勾配とも呼べる)が、割球の性質を規定し、後の発生に大きな影響を及ぼす例は両生類においても認められる。カエルの卵は卵割が進むと、卵黄を多く含む植物極の割球と、少ない動物極の割球に分かれる。このとき、卵細胞質に存在した極性はそれぞれの割球へと区画化されることになる。植物極の腹側の特定の割球に、ショウジョバエの時と同じく生殖細胞質が分配され、その細胞はやがて始原生殖細胞(primordial germ cell)になる。
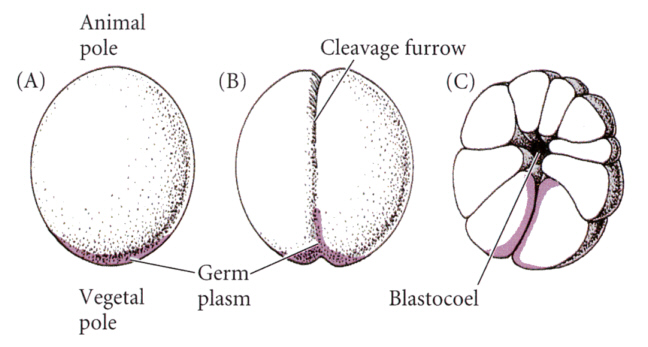
Developmental Biology, 6th ed. Scott Gilbert Sinauer Associates, Incより
2)始原生殖細胞
脊椎動物では、発生が進むと始原生殖細胞が大挙して移動し、やがて中胚葉起源の生殖隆起(genital ridge)へ落ち着く。マウスでは、7.5日目の胚の尿嚢付近に最初に観察され、その後まもなく尿嚢基部および尿嚢に隣接した卵黄嚢内に見いだされる。
細胞集団は後腸を経由して頭側方向へ移動し、背側腸管膜を背側へ移動して、生殖隆起内へ入る。ほとんどの始原生殖細胞は11日までに生殖隆起へ到着する。最初は10から100細胞だが、移動しながら増殖し、12日胚の生殖隆起内には2,500から5,000個の始原生殖細胞が存在する。
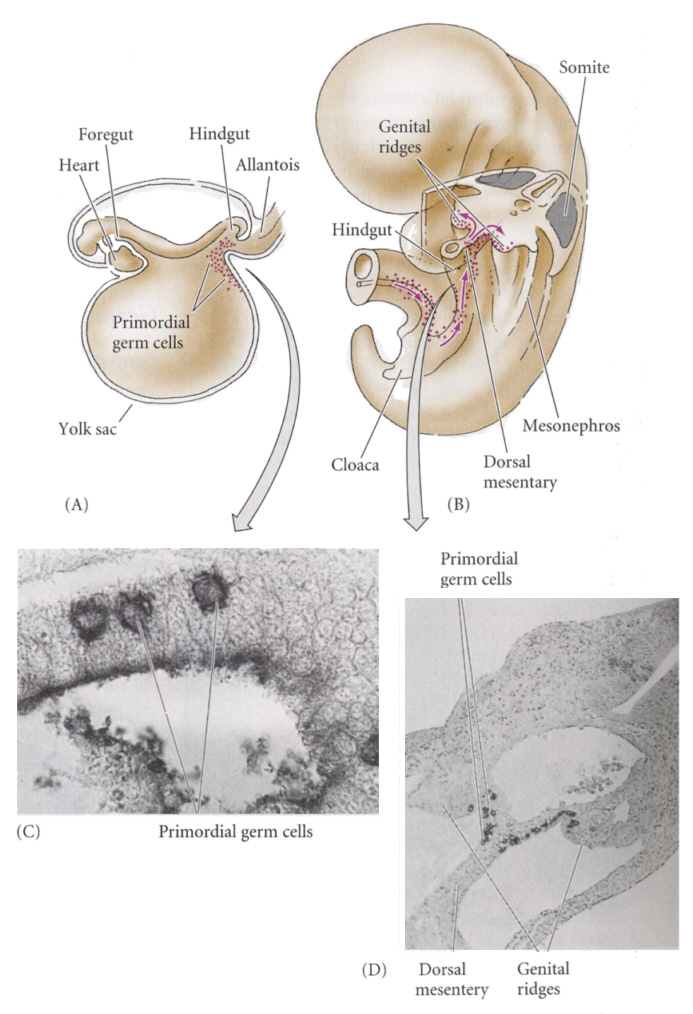
Developmental Biology, 6th ed. Scott Gilbert Sinauer Associates, Incより
鳥類では盤割をするが、頭突起の前方に生殖三日月環と呼ぶ部域が現われ、ここから始原生殖細胞が生じる。鳥類の始原生殖細胞は、両生類や哺乳類がアメーバ運動で移動するのと異なり、いったん血管内に入り、血流にのって生殖隆起に達し、ここで血管から出て生殖隆起に入る。
3)生殖器原基
このように、脊椎動物では「中身」の生殖細胞と「器」の生殖隆起とでは起源が異なっている。生殖隆起を構成する体腔上皮細胞が増殖、肥厚し、背方に向かって索状に伸び、そこへ生殖細胞が取り込まれる。
この構造を第一次性索と呼ぶ。この時点では、雄でも雌でも同じような生殖腺原基をもつ。
やがて雄では性索が網状組織を形成し、その最末端に薄い精巣網(rete testis)を作る。この生殖細胞が詰まった網状組織が、後に精細管(seminiferous tubule)になる。
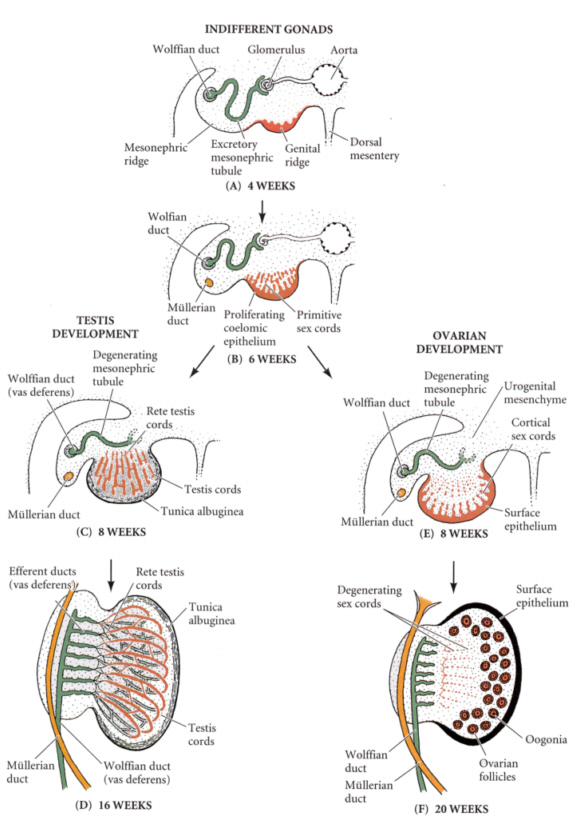
Developmental Biology, 6th ed. Scott Gilbert Sinauer Associates, Incより
一方雌では、生殖細胞は生殖腺の外表に近いものを残し退化し、性索も退化する。上皮はやがて伸張し、第二次性索(皮索)を形成する。皮索はやがて生殖細胞を1個含む細胞塊に分かれる。これが原始卵胞になる(卵形成参照)。
4)雌雄分化
雄になるか雌になるかは、まず染色体の組み合わせによって決まる。哺乳類では性染色体がXXなら雌となり、XYなら雄になる。しかしながら、本物の雄になるためにはまだまだたくさんの道のりがある。すなわち、生殖輸管の雌雄分化、外部生殖器の雌雄分化、脳の雌雄分化である。これらの過程が狂うと、染色体は男なのに、外見は女になってしまうことがありうる。
まだ未分化の状態では、雄にも雌にもウォルフ管とミューラー管が存在するが、やがて雄ではミューラー管が退化し、ウォルフ管が残り副精巣と輸精管となる。一方雌ではウォルフ管が退化し、ミューラー管が発達し、輸卵管、子宮、膣の一部となる。
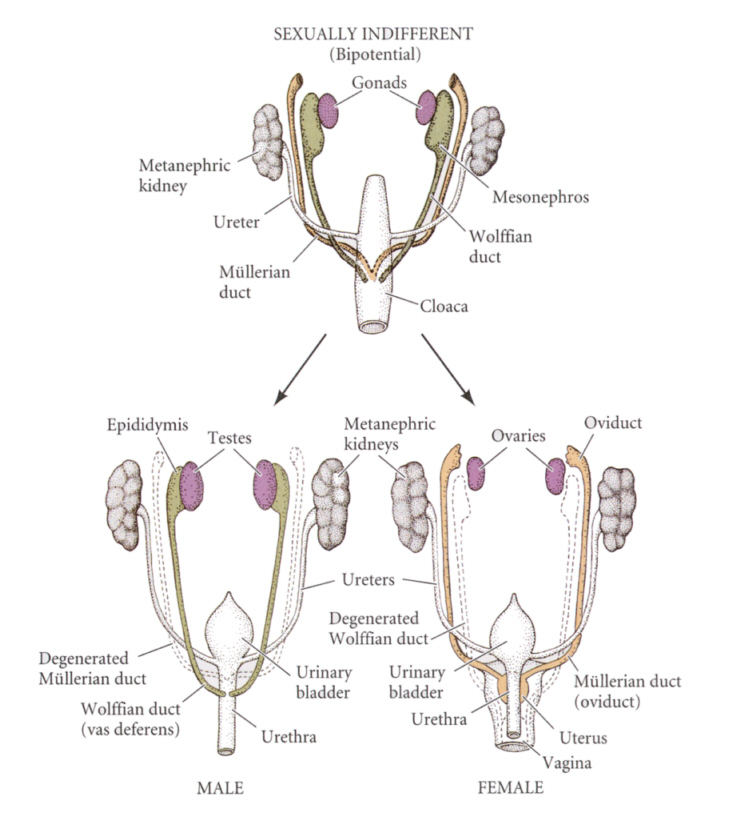
Developmental Biology, 6th ed. Scott Gilbert Sinauer Associates, Incより
遺伝的に雄のウサギの胎児(第19日目)で、ミューラー管もウォルフ管も性的に分化していないときに生殖腺を除去すると、ウォルフ管は退化し輸精管や前立腺は形成されない。しかし遺伝的に雄でありながら、ミューラー管は残り、輸卵管、子宮ができ、外部生殖器も雌型になる。遺伝的に雌の場合には生殖腺を除去しても雌型の生殖器官が発達する。つまり生殖腺がないと遺伝的に雄でも雌でも雌型の生殖器官が発達する(哺乳類の場合)。
胎児の精巣からはウォルフ管を発達させ、ミューラー管の発達を抑制する因子が出ていることを示している。ミューラー管の発達を抑制する因子は、560個のアミノ酸からなる糖タンパク質である抗ミューラー管ホルモン(AMH)であることが明らかになった。このホルモンは、セルトリ細胞から分泌される。一方、ウォルフ管は、精巣のライディッヒ細胞(間細胞)から分泌される、テストステロンによって維持される。テストステロンがないと、ウォルフ管はアポトーシスを起こして消失してしまう。
やがて、完成した生殖腺からは性ステロイドホルモンが分泌され、外部生殖器がそれぞれ雄のものと雌のものに分化する。そのため、性染色体がXYで男になるはずであっても、アンドロジェンの受容体がないと下の写真のように外部形態は女のものとなってしまう。

Developmental Biology, 6th ed. Scott Gilbert Sinauer Associates, Incより
生殖輸管と付属生殖器は、テストステロンによって男になるが、外部生殖器(ペニスや陰嚢)は、テストステロンが代謝されてできる5αジヒドロテストステロンによって男のものになる。したがって、この変換を触媒する酵素が遺伝的に欠損していると、外部生殖器は男のものにはならない。
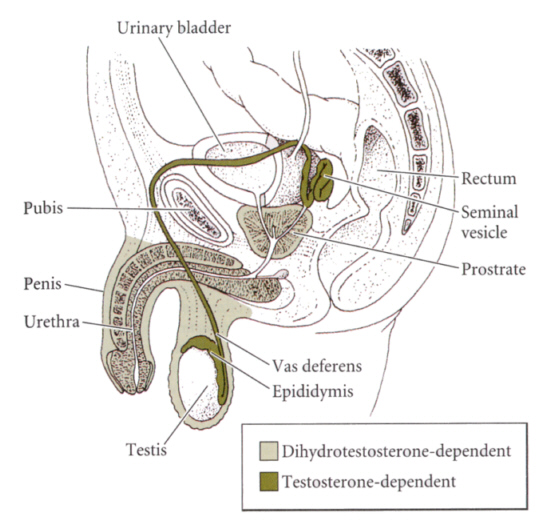
Developmental Biology, 6th ed. Scott Gilbert Sinauer Associates, Incより
それではXYだとどうして雄になるのだろうか。長い探索の末に、Y染色体にSry(sex-determining region of the Y)と呼ばれる遺伝子が存在することがわかった。この遺伝子はY染色体の短椀上に存在する。この遺伝子とプロモータ部位を含む14kbのDNAをマウスに導入した、いわゆるトランスジェニックマウス雌(染色体はXX)で、この遺伝子が発現し、精巣や雄の生殖輸管、外部生殖器が発達することが確認された。
しかしながら、性染色体以外にも性を決定するいくつかの遺伝子が存在するらしく、性の決定のメカニズムが完全には解明されたわけではない。
ページ先頭へ戻る
![]() この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
 ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。
ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。