第8章 DNAからタンパク質へ
1.DNAの発見と染色体地図
2.DNAが遺伝情報を担っていることの発見
3.DNAの構造
4.DNAからタンパク質へ
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
更新日:2003/05/07
これまではメンデルの要素と遺伝子あるいはDNAを、あまり厳密に区別をせずに使ってきた。また遺伝子が染色体に載っているとも言ってきた。第7章で染色体とDNAの構造の関係を述べたが、染色体に遺伝子が載っているというのはどういうことだろうか?
ちょうど今から50年前の1953年に、DNAの構造が明らかになった。ここではDNAの構造が明らかになるまでと、DNAと遺伝子について整理しながら理解していこう。
http://www.dnaftb.org/dnaftb/(とてもよい)
1)DNAの発見
物質としてのDNAが発見されたのは比較的古く、1869年のことである。ダーウィンの進化論の発表、メンデルの遺伝の実験の発表とほぼ同時期である。発見したのはFriedrich
Miescher(1844-1895)である。ミーシャーはスイスのバーゼルで生まれ、牧師になりたかったが、病理学教授であった父親の反対で医学の道に進んだ。聴覚障害のために基礎医学の研究をめざし、チュービンゲン大学のホッペザイラーのもとで研究をおこなった。
 ホッペザイラーは、細胞説にもとづき細胞の化学的な裏づけを得ようとしていた。ミーシャーに与えられたテーマは、白血球の細胞成分の化学的な研究であった。白血球を生体から多量に得るのは難しかったので、彼は膿に着目し、病院で多量にでるガーゼに付着した膿を集めた。この死んだ白血球の核から新しい物質としてC、H、O以外にリンと窒素を含むヌクレイン(現在のDNA)を抽出する。ミーシャーはヌクレインの研究を続け、酵母、腎臓、肝臓などにも同じ物質が含まれていることを明らかにする。後に膿の代わりにサケの精子を使い、多量のヌクレインが含まれることを見つけるとともに、精子細胞核中に特異的なタンパク質をも見つけた。
ホッペザイラーは、細胞説にもとづき細胞の化学的な裏づけを得ようとしていた。ミーシャーに与えられたテーマは、白血球の細胞成分の化学的な研究であった。白血球を生体から多量に得るのは難しかったので、彼は膿に着目し、病院で多量にでるガーゼに付着した膿を集めた。この死んだ白血球の核から新しい物質としてC、H、O以外にリンと窒素を含むヌクレイン(現在のDNA)を抽出する。ミーシャーはヌクレインの研究を続け、酵母、腎臓、肝臓などにも同じ物質が含まれていることを明らかにする。後に膿の代わりにサケの精子を使い、多量のヌクレインが含まれることを見つけるとともに、精子細胞核中に特異的なタンパク質をも見つけた。
ミーシャーはヌクレインの化学組成を明らかにしているが、ヌクレインが遺伝に関与する物質であるとは考えていなかった。1940年代までは、誰もが遺伝を担う物質はタンパク質であろうと考えていた。したがってヌクレインは機能不明な物質としてしばらくは日の目を見なかったのである。
2)染色体地図
 DNAの発見とは別に、染色体と遺伝子の関係がThomas
Hunt Morgan(1866-1945)の研究によって1920年代になってさらに明らかになった。
DNAの発見とは別に、染色体と遺伝子の関係がThomas
Hunt Morgan(1866-1945)の研究によって1920年代になってさらに明らかになった。
モーガンはショウジョウバエを使って、メンデルと同じような遺伝の実験をおこなった。有名な実験は赤眼のショウジョウバエと突然変異体として発見した白眼のショウジョウバエを使った伴性遺伝の実験である。これは第5章の伴性遺伝の例として述べた血友病の場合とまったく同じ形式の遺伝をする。
 一世代の時間が短いので、ショウジョウバエは遺伝の実験をおこなうのに便利だった。また突然変異体を比較的容易に作り出すことができる。モーガンらはこの利点を生かして、さらに二遺伝子雑種の実験をおこなった。この実験には体色が黒くなる突然変異体(b)と痕跡翅となる突然変異体(vg)を使った。メンデルの独立の法則に従うのならば、雑種第二代では、2つの遺伝子の分離比は9:3:3:1になるはずである。ところが結果はこれと大きくずれていた。そこで戻し交配(F1と劣性ホモを掛け合わせて、そのこどもの表現型の比を調べることにより、遺伝子型の比を求める方法)をおこなってみたところ、BVg:Bvg:bVg:bvgの比は965:206:185:944であった。
一世代の時間が短いので、ショウジョウバエは遺伝の実験をおこなうのに便利だった。また突然変異体を比較的容易に作り出すことができる。モーガンらはこの利点を生かして、さらに二遺伝子雑種の実験をおこなった。この実験には体色が黒くなる突然変異体(b)と痕跡翅となる突然変異体(vg)を使った。メンデルの独立の法則に従うのならば、雑種第二代では、2つの遺伝子の分離比は9:3:3:1になるはずである。ところが結果はこれと大きくずれていた。そこで戻し交配(F1と劣性ホモを掛け合わせて、そのこどもの表現型の比を調べることにより、遺伝子型の比を求める方法)をおこなってみたところ、BVg:Bvg:bVg:bvgの比は965:206:185:944であった。
この値から組み換え率を計算すると、(206+185)/(965+206+185+944)x100=17%となる。これはこの二つの遺伝子が同じ染色体上にあり、生殖細胞をつくる過程で染色体の交叉によって遺伝子の組換えが起こったと考えると説明がつく。
モーガンらはこのような組み換えを徹底的に調べ、組み換えがおこる連鎖群が4つあることを明らかにする。これはショウジョウバエの染色体(2n=8)の半数に等しい。さらに上のような組み換え率を計算し、それぞれの連鎖群内の、遺伝子座の相対的な位置関係を計算によって求めた。こうしてショウジョウバエの染色体地図がつくられ、遺伝子は染色体の上に直線状に並んで載っていることが明らかになった。
連鎖と組み換えに関しては、下記のサイトも参照してください。
http://www.tmd.ac.jp/artsci/biol/textintro/introtop.htm
http://biology.uky.edu/MIF/thm.html#anchor1116140(モーガンについて)
3)一遺伝子-一酵素説
エンドウの花の色やショウジョウバエの眼の色で示されたように、遺伝子は表現型を規定している。実際に遺伝子は花の色や眼の色と言う表現型をどのように規定しているのであろうか。この点を明確にしたのがビードルとテイタム(George
W. Beadle and Edward L. Tatum)で、1941年のことである。

 ビードル(左)は、最初はモーガンの研究室でショウジョウバエの眼の色に関する遺伝の研究をおこなった。1935年までに、赤い眼の色は、遺伝的に決められた一連の化学反応の結果、生じることを示唆する証拠を得ていた。その後、生化学者のテイタム(右)と共同研究をおこなうが、ショウジョウバエの眼の色では化学反応と遺伝子の関連を示すためにはあまりにも複雑すぎるので、ショウジョウバエからアカパンカビに実験材料を切り替えることにした。
ビードル(左)は、最初はモーガンの研究室でショウジョウバエの眼の色に関する遺伝の研究をおこなった。1935年までに、赤い眼の色は、遺伝的に決められた一連の化学反応の結果、生じることを示唆する証拠を得ていた。その後、生化学者のテイタム(右)と共同研究をおこなうが、ショウジョウバエの眼の色では化学反応と遺伝子の関連を示すためにはあまりにも複雑すぎるので、ショウジョウバエからアカパンカビに実験材料を切り替えることにした。
アカパンカビは、グルコース、無機塩類、ビオチン(ビタミンの一種)を含む最小培地で培養することができる。有性生殖によって胞子を作り、この胞子は無性生殖によってドンドン増えてコロニーを作る。好都合なことに胞子は半数体なので、突然変異の結果がそのまま表現型にあらわれる。
ビードルはまずアカパンカビにX線を照射して突然変異体をつくった。 こうして得た突然変異体の中に、栄養要求性の突然変異体があった。すなわち、最小培地では生育できず、培地に酵母の抽出物を加えると生育できるようになる変異体である。栄養要求性の突然変異体をさらに調べたところ、突然変異体の中には、1種類のアミノ酸を添加すれば生育できるものがあることが分かった。
こうして得た突然変異体の中に、栄養要求性の突然変異体があった。すなわち、最小培地では生育できず、培地に酵母の抽出物を加えると生育できるようになる変異体である。栄養要求性の突然変異体をさらに調べたところ、突然変異体の中には、1種類のアミノ酸を添加すれば生育できるものがあることが分かった。
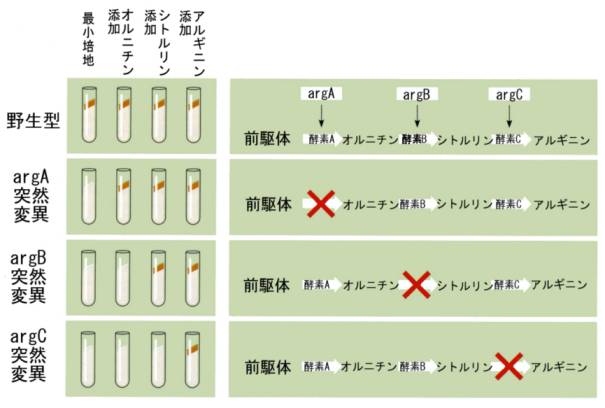
彼らはアルギニン要求性の突然変異体に注目して調べたところ、アルギニン要求性の突然変異体には3つの系統があることが分かった。これらの系統をargA、argB、argCと名づけることにしよう。こうして表現型は、眼の色のような目に見えるものから、栄養要求性という眼には見えないものに拡張されたのである。
野生型のアカパンカビは、もちろん最小培地で生育することができる。ところがargA突然変異体は最小培地では生育できず、オルニチンを加えた培地であれば生育することができた。また、argBはオルニチンを加えただけでは生育できず、シトルリンを加えたところ生育できた。3番目のargCはオルニチンでもシトルリンでもだめで、アルギニンを加えてはじめて生育することができた(1945年に発表)。
これらの結果を説明するためには、アルギニンがアカパンカビの中で生合成される経路(前駆物質 → オルニチン → シトルリン → アルギニン)があって、その各ステップを触媒する酵素が、argA、argB、argCという遺伝子によってコードされていると考えるとうまく説明ができる。
こうして、遺伝子は眼の色と言う漠然としたものではなく、酵素という実体のあるタンパク質をコードしていることが明確になったのである。
実験と結果 結果の解釈
この結果から、ビードルとテイタムは一遺伝子-一酵素説という仮説を提唱した。その後、この仮説は少し変更を受ける。酵素のなかには複数のポリペプチド鎖から構成されるものがあり、その場合は、遺伝子は一つではなく、複数になるからである。また遺伝子は酵素だけではなく、構造タンパク質をもコードしている。したがって現在では、一遺伝子-一ポリペプチド鎖と言うほうが正しい。
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/N/Neurospora.html
ページの先頭へ戻る
染色体はヌクレインすなわちDNAとタンパク質でできていることが分かり、染色体上に遺伝子が載っていることが明らかになったので、遺伝子の本体はDNAかタンパク質のどちらかだということになる。すでに述べたように、核酸は4種類のヌクレオチドからできているのに対して、タンパク質は20種類のアミノ酸からなり、より複雑な構造をとことができるので、タンパク質の方が遺伝情報を担うのにふさわしい、と漠然と考えられていた。
これに対して、DNAが遺伝情報を担っているのことを示唆する研究があらわれる。これらの研究は、エンドウやショウジョウバエよりも簡単な構造の細菌やウイルスを使っておこなわれた。
1)グリフィスの実験
まずイギリスのFrederick
Griffithが1928年に肺炎双球菌を(Streptococcus
pneumonia)使った実験をおこなった。肺炎双球菌には2系統あり、1つ目の系統は野生型で病原性があり、マウスでは致死性である。挟膜をもち、培地に撒いて培養すると、縁が滑らかなコロニーをつくる(S型、smooth)。もう一つの系統は突然変異体で病原性を失っており、挟膜がなく、培地に撒いて培養すると縁がギザギザなコロニーをつくる(R型、rough)。肺炎双球菌は煮沸によって殺すことができる。
グリフィスは、この2つの系統をマウスに注射して、S型では確かにマウスが死んでしまい、R型では死なないことを確かめた。次に、S型を加熱して殺してから注射すると、マウスは死なないことを確かめる。
ところが、病原性の無いR型に、加熱して殺したS型を混ぜてから注射すると、注射されたマウスは死亡した。死んだマウスの血液中からは培養すると縁が滑らかなコロニーを作る菌が得られた。
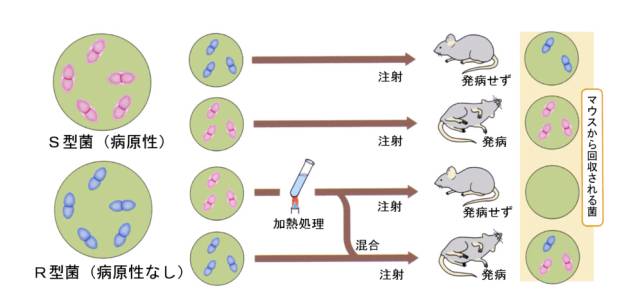
これらの実験結果は、R型の肺炎双球菌がS型の何らかの因子によって病原性を持つように形質が転換したことを示している。グリフィスはこれを形質転換因子と名づけたが、因子の本体については明らかにすることはできなかった。
2)アベリーの実験
グリフィスの実験を受けてアメリカのOswald
Theodore Averyらは、形質転換因子がどのような物質であるかの追求をおこなった。
 S型から抽出した形質転換因子を加えると形質転換が起こるのだから、抽出物中のいろいろな物質を順番に壊して形質転換が起こるかどうかを試してみればいい。そこでS型菌からの抽出物を遠心分離して、分子量の大きな分画を除いた上清で試みたところ、形質転換はおこった。そこで、上清をタンパク質分解酵素で処理したが形質転換は起こった。またRNA分解酵素でも影響はなかった。ところが、DNA分解酵素で処理すると形質転換は起こらなくなった。つまりDNAが形質転換因子だったのである(1944)。
S型から抽出した形質転換因子を加えると形質転換が起こるのだから、抽出物中のいろいろな物質を順番に壊して形質転換が起こるかどうかを試してみればいい。そこでS型菌からの抽出物を遠心分離して、分子量の大きな分画を除いた上清で試みたところ、形質転換はおこった。そこで、上清をタンパク質分解酵素で処理したが形質転換は起こった。またRNA分解酵素でも影響はなかった。ところが、DNA分解酵素で処理すると形質転換は起こらなくなった。つまりDNAが形質転換因子だったのである(1944)。
こうしてDNAが形質を転換する因子の本体であること、すなわち遺伝子の本体であることを強く示唆する結果が公表されたが、多くの人はまだ半信半疑だった。細菌やウイルスに遺伝子としてのDNAがあることさえも、必ずしも明確ではなかったからである。
3)バクテリオファージを使った実験
 ウイルスはDNAにタンパク質の衣をかぶせたようなもので、生物とも無生物ともいえる不思議な生き物である。ウイルスは自らタンパク質を合成できないので、細菌や他の生物の細胞内に入り込んで、その細胞のタンパク質合成工場を乗っ取ってタンパク質の衣をつくる。バクテリアを宿主とするものをバクテリオファージ(あるいは単にファージ)という。
ウイルスはDNAにタンパク質の衣をかぶせたようなもので、生物とも無生物ともいえる不思議な生き物である。ウイルスは自らタンパク質を合成できないので、細菌や他の生物の細胞内に入り込んで、その細胞のタンパク質合成工場を乗っ取ってタンパク質の衣をつくる。バクテリアを宿主とするものをバクテリオファージ(あるいは単にファージ)という。
物理学から転進したMax
Delbruckは1937年にアメリカに渡り、ルリアやハーシーとファージ研究グループを立ち上げ、細菌とファージの分子遺伝学の基礎を築いた。彼らは、大腸菌と大腸菌を宿主とするT系バクテリオファージに研究を集中するように提案して、この分野の研究を推し進めた。
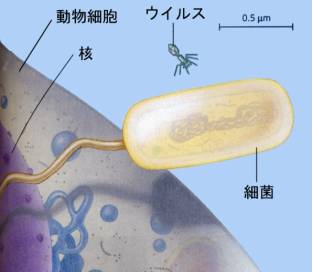

T系バクテリオファージは月着陸船のような構造をしていて、大腸菌に取り付くと中身を大腸菌の中に注入し、やがて大腸菌の中で月着陸船のようなタンパク質の衣とDNAを複製して増殖し、大腸菌を破って飛び出してくる。
それではファージは、大腸菌の中でDNAを使って自分と同じファージをたくさん作りだしているのだろうか。あるいはタンパク質を使っているのだろうか。この点を明らかにしたのがAlfred
Day HersheyとMartha
Chaseで、彼らはblender
experimentという巧みな実験系を組んでこれを証明した。
タンパク質を構成するアミノ酸はCHON以外にメチオニン(アミノ酸の一つ)ではSを含む。一方の核酸の構成要素であるヌクレオチドでは、CHON以外にPを含む。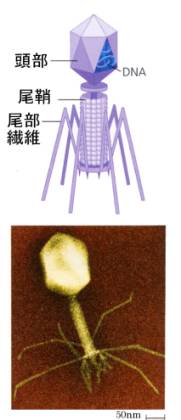
そこで、一方のバッチでは放射性Sで標識したメチオニンを含む培地で大腸菌を飼ってファージに感染させ、外皮タンパク質を放射性Sで標識する。もう一つのバッチでは、放射性Pで標識したヌクレオチドを含む培地で大腸菌を飼い、ファージに感染させてDNAを放射性Pで標識する。
この2種類の標識をしたファージとブレンダーを使って、彼らは次のような実験をおこなった。
一定時間培養した後に、ブレンダーで攪拌してファージを大腸菌から離し、遠心して上清と沈殿したペレット(この中に大腸菌の菌体が含まれている)の放射能を調べた。その結果、タンパク質を標識した場合は上清に放射能が現れ、DNAを標識した場合はペレットに放射能が現れることが示された。大腸菌に入るのはDNAだけだったのである。

こうして、T系ファージはDNAを菌体内に注入し、タンパク質の衣は菌体内には入らないことが明らかにされた。下の写真は、T2ファージが大腸菌に付着してDNAを注入しているところを撮影した電子顕微鏡写真に着色したものである。
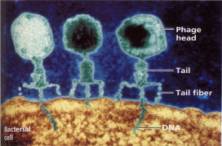
こうして、遺伝子の本体はDNAであることが確定したのである。
ページの先頭へ戻る
1)DNAの化学的性質の研究
1920年代に生化学者のLeveneがDNAの化学的組成について研究をおこい、DNAは4種類の窒素を含む塩基、すなわちシトシン(C)、チミン(T)、アデニン(A)、グアニン(G)、デオキシリボースという五炭糖とリン酸で構成されていることを発見する。レビンは、DNAの単位はヌクレオチドで、デオキシリボースに塩基とリン酸が結合していると考えた。しかしながらレビンは、4種類の塩基の比は等しく、DNAの構造として、ヌクレオチド4つを単位としたテトラマーが繰り返し結合しているという、今となっては誤った結論を下してしまった。
 1949年になってErwin
ChargaffはDNAの塩基の組成を調べ、4種の塩基の比は等しくないが、AとTおよびGとCの量が等しいと言う関係があることを見つけ、したがってプリン塩基(A+G)=ピリミジン塩基(T+C)という関係があることを明確にした。このことは次に述べるワトソンとクリックがDNAのモデルを作り上げるのに大きな手がかりとなった。
1949年になってErwin
ChargaffはDNAの塩基の組成を調べ、4種の塩基の比は等しくないが、AとTおよびGとCの量が等しいと言う関係があることを見つけ、したがってプリン塩基(A+G)=ピリミジン塩基(T+C)という関係があることを明確にした。このことは次に述べるワトソンとクリックがDNAのモデルを作り上げるのに大きな手がかりとなった。
ここで、第2章で学んだDNAの構成単位であるヌクレオチドについて復習しておこう。
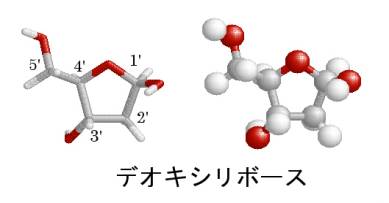
ヌクレオチドはレビンが考えたように、デオキシリボースという五炭糖に、リン酸と塩基が結合した分子である。五炭糖であるデオキシリボースの炭素を区別するために、右の炭素から順番に時計回りに1’から5’の番号を付ける。塩基は1’の炭素に結合し、リン酸は5’の炭素に結合している。3’の炭素には水酸基がつくが、2’の炭素には水酸基はない。
塩基にはアデニン、チミン、グアニン、シトシンの4種があるので、DNAを構成するヌクレオシドにもそれに従って、アデノシン、チミジン、グアノシン、シチジンの4種類がある。
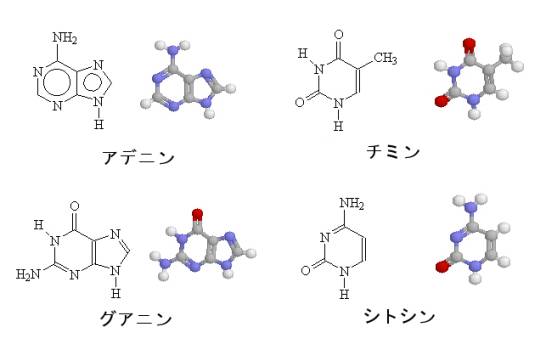
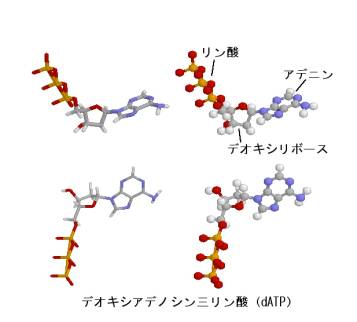
塩基以外の構造は4種のヌクレオシドでまったく同じである。ヌクレオシドにリン酸がついたものがヌクレオチドである。上の図は、リン酸が3つ5’についたデオキシアデノシン三リン酸である。他のヌクレオチドは、分子の右側に描かれた塩基を、それぞれチミン、グアニン、シトシンに変えたものとなる。
DNAは、この4種のヌクレオチドが直線状につながったものであることはわかった。それがどのようなつながり方をしているかは、まだわからなかった。
2)ワトソン・クリックのモデル

 1951年にアメリカで学位を取ったばかりのJames
D. Watsonが、イギリスでFrancis Harry Compton
Crickと出会った。これがDNA構造の解明への第一歩だった。
1951年にアメリカで学位を取ったばかりのJames
D. Watsonが、イギリスでFrancis Harry Compton
Crickと出会った。これがDNA構造の解明への第一歩だった。
ワトソンはファージ研究グループのところで述べたルリアのもとで博士号を取得した後、タンパク質の研究のために留学したコペンハーゲンから、ケンブリッジ大学のキャベンディッシュ研究所にやってきたのである。彼はシュレーディンガーの『生命とは何か』を読んで、生命の謎を解くのは遺伝子の解明だと心に期するものがあった。ここで、PhD取得のために研究所で研究をしていたクリックと同室になる。
クリックは物理学を学んだ後、大学院に進むが戦争で中断され、海軍省で働くことになる。戦後、新しい道をと生物学の分野に移り、X線回折で有名なローレンス・ブラッグ卿が開設した研究所のMax
Perutzの研究室にやってきたのである。
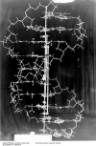 タンパク質ではなくDNAの話ですっかり意気投合した二人は、DNAの構造を解明するために、部屋の中に大きな模型を組んでジグソーパズルのような謎解きを始めることになる。
タンパク質ではなくDNAの話ですっかり意気投合した二人は、DNAの構造を解明するために、部屋の中に大きな模型を組んでジグソーパズルのような謎解きを始めることになる。
このときアメリカではタンパク質の二次構造であるαヘリックス構造を解明してノーベル賞をすでに授賞していたLinus
Paulingが、2つ目のノーベル賞を目指して、やはりDNAの構造解明の研究を始めていた。
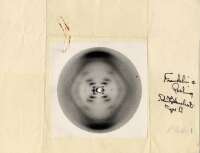 二人が有利だったのは、X線回折のデータが得られたことであった。
Maurice Wilkinsのもとで研究をしていたRosalind
Franklinが美しい回折像を提供した。
二人が有利だったのは、X線回折のデータが得られたことであった。
Maurice Wilkinsのもとで研究をしていたRosalind
Franklinが美しい回折像を提供した。
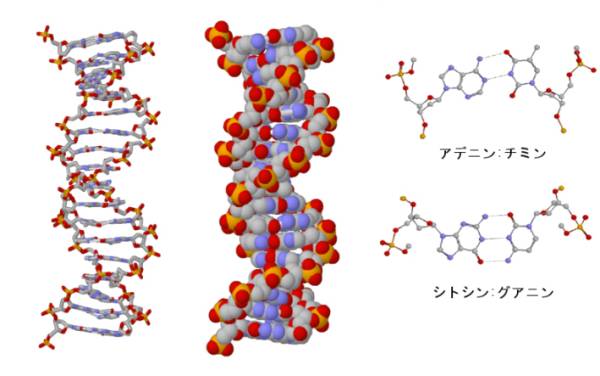
ある朝、ワトソンは、A:TおよびG:Cが水素結合をつくると考えるとピッタリと収まることに気が付き、これを聞いたクリックは、αヘリックスでは側鎖がラセンから外に突き出ているが、DNAでは塩基が内側を向いて二重ラセン構造をとれば、シャルガフの経験則とX線回折像を説明でき、ジグソーパズルがピッタリと収まることをすぐに理解した。こうしてDNAの分子模型はこの世に現れたのである。1953年2月28日土曜日のことであった。
わずか1ページの短い論文はNatureに投稿されて掲載される。このモデルがすぐに受け入れられたのは、A:TおよびG:Cがそれぞれ2本及び3本の水素結合で結合し、それ以外の組み合わせでは結合できないという点である(相補性、complementary)。これによって、細胞分裂のときに染色体が複製されて同じ物が娘細胞に分配されるという現象を、分子のレベルでみごとに説明できたからである。
http://biocrs.biomed.brown.edu/Books/Chapters/Ch%208/DH-Paper.html(原著論文)
http://molvis.sdsc.edu/dna/fs_pairs.htm(Chimeが必要)
http://www.chm.bris.ac.uk/motm/dna/dnac.htm
 1962年にワトソンとクリックはウィルキンスとともにノーベル医学生理学賞を授賞する。この時、フランクリンはこの世にはいなかった。1958年にガンでなくなっていたのである。ノーベル賞は死者には与えられない。またシャルガフも「賞は3人まで」の壁のために受賞を逸した。ちなみに、ポーリングは同じ年にノーベル平和賞を受賞する。また、ペルツもJohn
Cowdery Kendrewとともに、X線回折によるミオグロビン構造解明によりノーベル化学賞を受賞する。
1962年にワトソンとクリックはウィルキンスとともにノーベル医学生理学賞を授賞する。この時、フランクリンはこの世にはいなかった。1958年にガンでなくなっていたのである。ノーベル賞は死者には与えられない。またシャルガフも「賞は3人まで」の壁のために受賞を逸した。ちなみに、ポーリングは同じ年にノーベル平和賞を受賞する。また、ペルツもJohn
Cowdery Kendrewとともに、X線回折によるミオグロビン構造解明によりノーベル化学賞を受賞する。
ポーリングとのDNAの構造解明競争の内幕を、後にワトソンは『二重ラセン』という本にして出版する(1968)。この本は、それまでのこの種の本とは異なり、科学が人間の活動だということをわからせる異色の本だった。ありきたりの自伝や偉人伝とは異なり、かなりきわどい内容を含んでいたこともあって、今でも版を重ねている。
http://osulibrary.orst.edu/specialcollections/coll/pauling/dna/index.html(ポーリングとの競争について)
ページの先頭へ戻る
こうして、DNAが遺伝子であり、その情報を使ってタンパク質を作り出していることが明らかになった。
次に問題になるのは、4種しかないDNAのヌクレオチドをどのように使って遺伝の情報としているか、また、DNAからどのようにしてタンパク質が実際に作られるのか、という問題だった。
すでにお話したように、DNA(の一方の鎖)もポリペプチド鎖のどちらも、それぞれ4種のヌクレオチドと20種のアミノ酸が直線状に連結したポリマーである。しかも、DNAの方は5’→3’、ポリペプチド鎖の方はN端→C端という方向性がある。2つの間の対応を取ることは容易であるように思われる。しかしながら、DNAは核の中にあり核から外に出ることはなく、タンパク質はサイトゾールで合成される。どのようにして両者が結びつくのだろうか。また4つと20ではどうしても数があわない。
 これらの難問に対して、クリックは新しい実験を促すようないくつもの仮説(セントラルドグマ、アダプター仮説など)を提出した。
これらの難問に対して、クリックは新しい実験を促すようないくつもの仮説(セントラルドグマ、アダプター仮説など)を提出した。
まず、塩基1個にアミノ酸に1個では足りないのだから、塩基複数個で対応させればいいということになる。2個では16通りでまだ足りないので、3個64の組み合わせが妥当であろうと考えた。余った部分は重複していると考えればいい。また、DNAとポリペプチド鎖をつなぐためにRNAを間において、情報はDNA→RNA→ポリペプチドというように流れると考え、アミノ酸を合成の場につれてくるアダプターを別に考えればいいという仮説を立てた。 これらの仮説は、後に実験によって証明される。
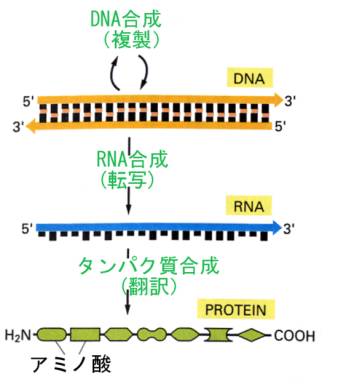
現在では、DNAからタンパク質への情報の流れは次のように考えられている。
DNAはまず、2本の鎖のうち、片方の鎖を鋳型として、相補性を利用してDNAの塩基配列をRNAに写し取る。この過程を転写(transcription)と言い、転写された一定の長さのRNAをメッセンジャーRNA(略してmRNA)と言う。
次に、mRNAの塩基の配列3つづつ(これをコドンと言う)に対応する運搬RNA(略してtRNA、クリックのアダプター)がmRNAの塩基配列にしたがって順番に並び、それぞれのtRNAに結合したアミノ酸がペプチド結合で結合すれば、DNAの塩基配列の情報に従ったアミノ酸配列のポリペプチド鎖ができあがる。
塩基の配列が遺伝の暗号であることも実際に確かめられた(1961-65)。合成したポリウリジンをin
vitroのタンパク合成系に入れると、ポリフェニルアラニンが合成されたのである。この実験を最初として次々の合成実験がおこなわれ、64種類の塩基の組み合わせに、それぞれ対応するアミノ酸が決められた。
次の暗号表は、こうして決められたDNAの塩基3つとアミノ酸の対応表である。
|
|
2番目の塩基
|
|
|
T
|
C
|
A
|
G
|
|
1
番
目
の
塩
基
|
T
|
Phe
|
Ser
|
Tyr
|
Cys
|
T
|
3
番
目
の
塩
基
|
|
Phe
|
Ser
|
Tyr
|
Cys
|
C
|
|
Leu
|
Ser
|
Stop
|
Stop
|
A
|
|
Leu
|
Ser
|
Stop
|
Trp
|
G
|
|
C
|
Leu
|
Pro
|
His
|
Arg
|
T
|
|
Leu
|
Pro
|
His
|
Arg
|
C
|
|
Leu
|
Pro
|
Gln
|
Arg
|
A
|
|
Leu
|
Pro
|
Gln
|
Arg
|
G
|
|
A
|
Ile
|
Thr
|
Asn
|
Ser
|
T
|
|
Ile
|
Thr
|
Asn
|
Ser
|
C
|
|
Ile
|
Thr
|
Lys
|
Arg
|
A
|
|
Met
|
Thr
|
Lys
|
Arg
|
G
|
|
G
|
Val
|
Ala
|
Asp
|
Gly
|
T
|
|
Val
|
Ala
|
Asp
|
Gly
|
C
|
|
Val
|
Ala
|
Glu
|
Gly
|
A
|
|
Val
|
Ala
|
Glu
|
Gly
|
G
|
Phe:フェニルアラニン、Leu:ロイシン、Ile:イソロイシン、Met:メチオニン、Val:バリン、Ser:セリン、Pro:プロリン、Thr:トレオニン、Ala:アラニン、Tyr:チロシン、His:ヒスチジン、Gln:グルタミン、Asn:アスパラギン、Lys:リシン、Asp:アスパラギン酸、Glu:グルタミン酸、Cys:システイン、Trp:トリプトファン、Arg:アルギニン、Ser:セリン、Gly:グリシン。なお、Metは開始コドンにもなり、Stopは終止コドンをあらわす。
こうして、染色体を構成しているタンパク質とDNAのうちDNAに、遺伝情報が塩基の配列というかたちで書き込まれていることが明らかになった。塩基4文字のうちの3つの組み合わせ(コドン)がアミノ酸を指定(コード)していたのである。遺伝子はポリペプチド鎖をコードする塩基配列で、これが染色体を構成するDNA分子上に線状に並んで載っているのである。
遺伝子は、対になった染色体(2n)にペアで存在し、細胞分裂によって誤りなく娘細胞に分配される。生殖細胞を作るときには半分(n)になる。こうして代々遺伝子は伝えられていくが、何らかの原因で塩基の文字が変わればアミノ酸も変わってしまい、タンパク質の構造も変わってしまう。タンパク質の構造が変わったために機能を失うばあもあるし、ほとんど影響が出ない場合もある。これが突然変異(mutation)である。こうした突然変異が個体群の変異の原因であり、自然選択を受ける対象となる。
 この章のpdfファイルをダウンロードするには、左の「Adobe」のアイコンを右クリックして、ファイルを保存を選んで、自分のパソコンにダウンロードしてください。
この章のpdfファイルをダウンロードするには、左の「Adobe」のアイコンを右クリックして、ファイルを保存を選んで、自分のパソコンにダウンロードしてください。
 ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。
ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。
ページの先頭へ戻る
生物学基礎トップページへ戻る