1.DNAの複製
2.細胞周期
3.体細胞分裂の過程
4.減数分裂の過程
5.細胞周期の調節
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください) 更新日:2003/05/21
今まで、あまり説明を加えず細胞分裂という言葉を使い、細胞分裂によって染色体は娘細胞に間違いなく同じ数だけ受け渡される、と述べてきた。ここでは、もう少し細胞分裂について調べてみよう。
細胞分裂(cell division)は、核の分裂と細胞質の分裂の2段階に分けられる。真核生物では、核が分裂するときには染色質が糸状の染色体となって分裂が進行するので、有糸分裂(mitosis)という。一方、生殖細胞を作るときには染色体の数を半減させるので、このような分裂の過程を減数分裂(meiosis)という。ここでは、用語の混乱を避けるために、母細胞が自分と同じ2つの娘細胞をつくる過程を体細胞分裂とし、生殖細胞をつくる過程を減数分裂と呼ぶことにする。
体細胞分裂の過程では、次の3つのことがおこる必要がある。
1)染色体を構成する遺伝情報の源であるDNAを正確にコピーして2倍にする必要がある。この過程をDNAの複製と呼ぶ。それから染色体を複製する。
2)ミトコンドリアのような細胞小器官も、それぞれの娘細胞に十分な量を均等に分配する必要がある。
3)細胞質を2つの娘細胞に分離する必要がある。
それぞれの過程がどのように進行していくか、順番に見ていこう。
http://www.emc.maricopa.edu/faculty/farabee/BIOBK/BioBookmito.html#Table%20of%20Contents
1)メセルセンとスタールの実験
1953年にワトソンとクリックがDNAの構造のモデルを短い論文で提出したときに、染色体が複製するときに必然的に必要なDNAの複製は、一方の鎖それぞれを鋳型にして新しい鎖が複製されて、2本の二本鎖DNAができるだろうと予想した。このような複製の仕方を半保存的複製(semi-conservative
replication)と呼んでいる。
この論文を見てMatthew MeselsonとFranklin Stahlはすぐに、これが実際に起こっていることかどうか実証する必要があると考えた。複製の方法には、半保存的複製以外に、保存的複製(もとの二本鎖DNAをそのまま鋳型にして、新しく二本鎖DNAが複製される)とランダム分断(日本鎖DNAを分断して複製し、再びつなぎ合わせる)が理論的にはありうると考えられるからである。
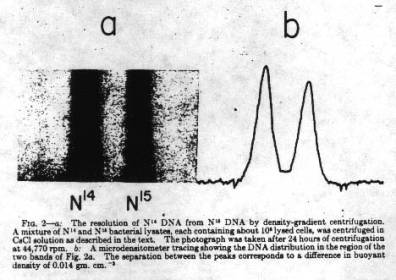
どれが正しかを証明するために、メセルセンとスタールはうまい方法を考え出す。DNAの重さの差を利用する方法である。CsClの濃い溶液を沈降セルに入れて超遠心機を使って大きなG(10,000g)をかけて遠心すると、セルの上から下に密度勾配ができる。溶液に重さ(密度)の異なる高分子を入れておくと、高分子はCsClの同じ密度のところに集まってくる。密度が異なる2つの高分子なら、2本のバンドができるはずである。彼らはまずこの方法で、実際にDNAが特定のバンドを作るかどうかを確かめた。
遠心時間が長くなると、間違いなく特定の場所にバンドができるのを確認した後、次にDNAに重さの差を出すために、14NH4OH のかわりに15NH4OHを培地に入れて何代も大腸菌を飼育した。こうすると大腸菌は塩基の材料として重たいNを使わざるを得ないのでDNAが重くなるのである。本当に重くなったかどうかも彼らは確かめる(下の原著の図参照)。
こうして重たい培地で飼育した大腸菌を、今度はふつうの14NH4OHで飼育し、最初の世代、次の世代、さらに次の世代と時間を追って大腸菌を集め、そこからDNAを集めて同じように密度勾配遠心法をおこなった。
第一世代の大腸菌から集めたDNAは1本のバンドになるが、その密度は第0世代のバンドよりも軽く、ちょうど重たいDNAと軽いDNAの中間であることを示していた。第二世代になるとバンドは2本に分かれ、1本は第一世代と同じ重さ、もう1本はさらに軽いバンドであった。次の図はそれを模式的に表わしてある。
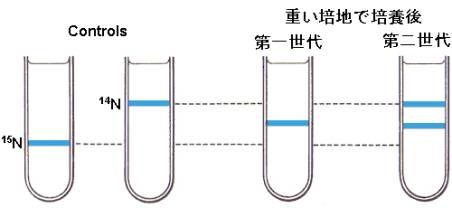
この実験は、明らかに新しく複製された二本鎖DNAの半分が軽いNの鎖で、もう一本が元の重いNのDNAであることを示している。こうして複製は予想通り、半保存的な様式でおこなわれることが実証されたのである。
大腸菌は原核生物であるが、その後、真核生物のDNAの複製も半保存的に起こっていることが確かめられる。真核生物のばあいは、一本のDNAは一本の染色体の構成単位なので、染色体の複製も半保存的に起こることになる。
彼らの工夫した密度勾配遠心法は、分子生物学の標準的な手法として、しばらくは盛んに使われるようになる。
http://www.blc.arizona.edu/marty/411/Modules/msexp.html(メゼルセンとスタールの実験)2)DNA複製の過程
DNAの複製は半保存的におこることが証明されたからといって、複製の詳しい過程が明らかになったのではない。複製がどのようにおこるかが明らかになるためには、さらに多くの研究者によるたくさんの研究が必要だった。複製の過程は生命現象の根幹部分なので、きわめて厳密に制御されている複雑な過程である。現在でも、複製の過程は完全に解明されているわけではない。ここでは基本的な部分だけをおさえておこう。
DNAからRNAへの転写にはRNAポリメラーゼが必要だったように、DNAの複製にはDNAポリメラーゼという酵素が必要である。ところが事はそう簡単ではない。転写の場合は片方のDNA鎖を鋳型として、相補性の規則にしたがって一本鎖のRNAを作ればよかったが、DNAの場合は両方の鎖をそれぞれ鋳型として、2本のDNA鎖を作らなければならない。ヌクレオチドから核酸を作るときには、かならず5’→3’の方向に合成が進むという制約があるので、二本鎖DNAの端にある両方の鎖に、いっぺんにヌクレオチドを付加して、鎖を伸ばしていくことはできない。ヌクレオチドの伸長は、次の図にあるように、原料となるヌクレオシド三リン酸のリン酸が2つ取れて、3’のOHとの間にエステル結合を作っていく過程である。
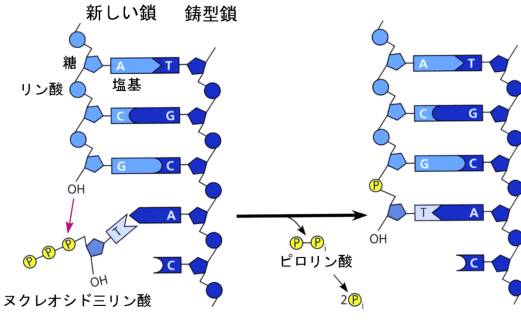
上の図では、左側の鎖の下端で下向きに5’→3’に伸びているが、右側の鎖では上端で反対方向(上向き)に伸びていくことになる。
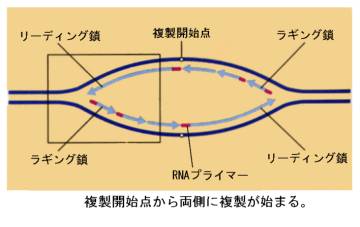
実際に複製が始まるためには、DNAは巻きついていたヒストンから離れて裸になり、さらにDNA二本鎖がほどけて複製が開始する。複製が始まるこのような場所を複製開始点という。複製開始点でDNAがほどけると、そこにDNAポリメラーゼが付着しDNAの複製がおこるのだが、方向が逆になるので2つのポリメラーゼがほどけた二本鎖DNAのそれぞれに付着して、DNAの二本鎖を押し広げる。そのため、複製開始点ではDNA鎖がY字状になるので、これを複製フォークと呼んでいる。
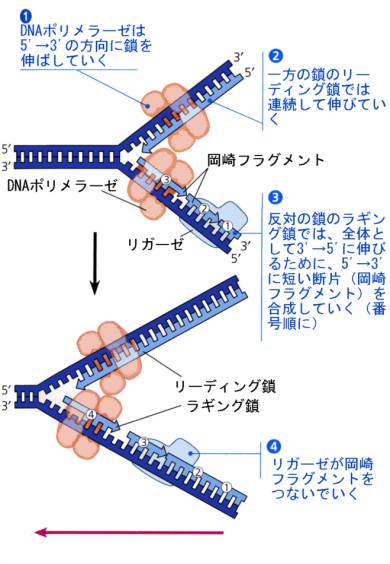
複製フォークの一方の鎖では鋳型鎖が3’→5’なので、DNAポリメラーゼはそのまま鎖を5’→3’に伸ばしていくことができる。順調に伸びていくこの鎖をリーディング鎖と呼ぶ。
もう一方の鎖であるラギング鎖は鋳型鎖が5’→3’なので、DNAポリメラーゼは3’→5’には合成できない。この点を細胞はどうやって解決しているかが謎であった。岡崎令次は、まずは短い断片として合成して、後でつなぎ合わせているのだろうと予想して、実験を行い、これを実証するデータを得て発表した(1966)。最初は受け入れられなかったがこれが正しいことが分かり、現在ではラギング鎖で最初に合成される短い断片をOkazaki fragmentと呼んでいる。
http://www.dent.tokushima-u.ac.jp/seiri/kana/notes/ROfragment.html(岡崎令次の実験)
現在では、下の図のような機構で複製が起こると考えられている。リーディング鎖では、DNAポリメラーゼが鋳型鎖に相補的なヌクレオチドを連続的につないでいく。反対側のラギング鎖では、別のDNAポリメラーゼがまず①をつくり、ある長さ合成されると複製開始点へ戻って②を合成し、また戻って③を合成するというようにDNAフラグメントを合成し、この断片をリガーゼがつないでいく。こうして、もとのDNAと全く同じ2本の娘DNAができあがる。
http://www.brh.co.jp/s_library/j_site/scientistweb/no32/
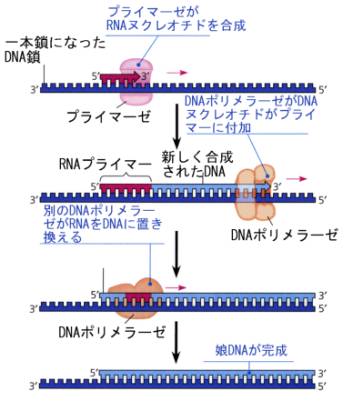
DNAポリメラーゼは一本鎖になった鋳型鎖にいきなりDNAヌクレオチドを合成していくことはできない。そのため、最初はRNAヌクレオチドがプライマーゼによって数個付加されてプライマーとなり、これを手がかりとしてDNAヌクレオチド付加されて伸びていくことが分かっている。
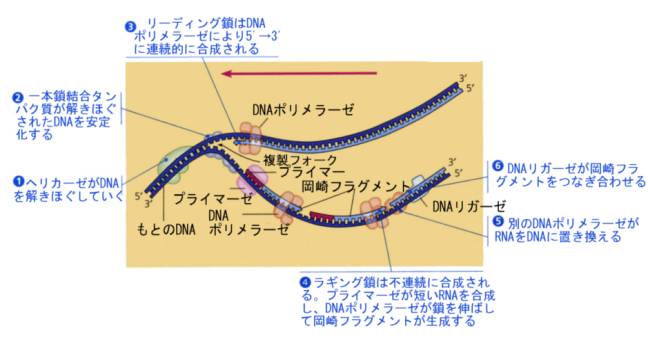
複製の過程には、上の図にあるように、この他にも多くの酵素が関与するが、ここでは省略する。
http://www.nig.ac.jp/museum/dataroom/replication/(複製の全体像、よくできている)
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/D/DNAReplication.html
上に述べたDNAの複製はいつもおこっているわけではない。細胞が分裂をしてその数を増やしていく時でも無秩序に増えていくのではなく、DNAの複製の項で述べたのと同じように、そのタイミングは厳密に制御されている。
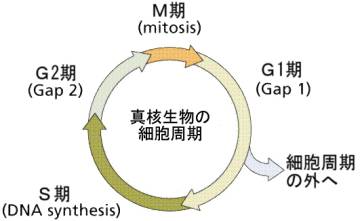
DNAの複製がおこる時期をS(synthesis)期といい、実際に染色体が現れて有糸分裂がおこる時期をM(mitosis)期という。M期とS期の間にあるギャップ期間をG1期、S期とM期の間をG2期という。細胞が数を増やしていく時にくり返す、このような周期を、細胞周期(cell cycle)という。M期以外の、G1、S、G2を間期(interphase)と呼ぶことがある。
植物の成長点とか上皮組織の基底部にある細胞のように、細胞分裂をくり返している細胞では、上に述べた細胞周期をくり返しているが、多くの細胞では、G1期で細胞周期の外へ出る。この状態をG0という。G0期の細胞でも、必要があると細胞周期の戻ることができる。たとえば肝臓の細胞はふだんはG0の状態にいるが、一部が切除されるとG0から細胞周期へ復帰し、細胞分裂をくり返して細胞の数を増やし、肝臓を元の大きさに戻す(肝再生)。G0期の細胞を細胞終期に戻すように働きかけるのは、成長因子やホルモンである。神経細胞のように細胞周期に戻れない細胞もある。
S期は2から4時間ほど続きG2期へ移行する。G2も2から4時間続いた後、M期へ移行し、体細胞分裂(1から2時間)がおこる。G0期は12時間から3日間続く。
http://www.cellsalive.com/cell_cycle.htm(細胞周期のアニメ)
体細胞分裂の過程のうちで、眼に見える形で核が分裂して、細胞が2つに分かれる過程をM期と呼んだが、M期はさらに前期(prophase)、中期(metaphase)、後期(anaphase)、終期(telophase)、細胞質分裂(cytokinesis)に分けることができる。もちろん本来は動的な過程なので、画然と区別できるわけではないが、便宜上このように分けて名前をつけている。
1)前期
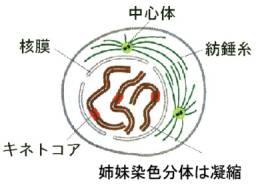
2)中期
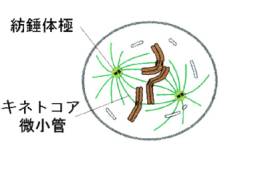
核膜の消滅が、前期前半の始まりのしるしである(prometaphase)。セントロメアに特殊なタンパク質が付着しキネトコアを形成し、ここへ両極からの微小管が向かい合って結合する(キネトコア微小管)。微小管の働きによって染色体が移動を始める。キネトコアと結合しなかった微小管同士は細胞の中央部でお互いに重なって結合し(極微小管)、全体として紡錘体(spindle)を形成する。
微小管の働きによって、すべての染色体が細胞中央部の一つの面(赤道面)に配列する(前期の後半)。こうしてすべての染色体が赤道面に並ぶことにより、染色分体が両極に移動して平等に2つに分かれることを保障する。
3)後期
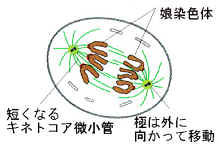
微小管の働きによって、染色分体は2つに引き離されて両極に移動し始める。両極からたキネトコアキネトコア微小管の短縮と、極微小管の相互作用による極を引き離す力によって、染色体はしだいに両極に移動する。
4)終期
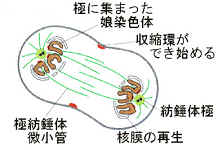
染色分体は両極に達し、染色分体を囲むように、まわりに核膜が形成され始める。染色体は染色質として核内に分散し始め、光学顕微鏡では見えなくなる。紡錘体が消失し、細胞質分裂が始める。
5)細胞質分裂
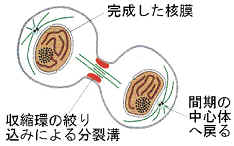
動物細胞では、赤道面の細胞表面に帯状にアクチンフィラメントが集まってくる(収縮環)。やがて、このアクチンフィラメントは収縮して細胞質を絞りこみ、2つの娘細胞にくびり切る。この過程で細胞小器官は2つの娘細胞に分配される。植物細胞では細胞壁があるために絞り込むことはできず、細胞板が形成されて細胞が2つに分かれる。
上の模式図は動物細胞の場合だが、次の写真は、植物細胞のの前期、中期、後期、終期の様子である。
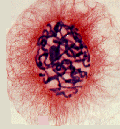
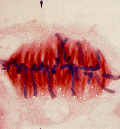


http://www.uoguelph.ca/zoology/devobio/210labs/mitosis1.html
http://www.cellsalive.com/mitosis.htm
S期における正確なDNAの複製と、複製したDNAをもとに染色体を複製して正確に2つの娘細胞に分け、さらに細胞小器官を分配することによって、母細胞からまったく同じ遺伝的組成を持った2つの娘細胞が作られる。我々の体は、このような過程によって、全く同じ遺伝的組成を持ったたくさんの細胞を受精卵から増やして、個体を形成しているのである。
1)減数分裂とは
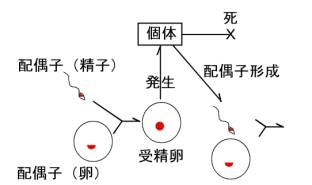
体細胞分裂は右の図で受精卵から個体が発生する過程と、個体を維持するために必要に応じて細胞の数を増やす過程で見られる。一方、減数分裂は、動物では、配偶子(精子と卵)の形成されるときに見られる。卵と精子は染色体の数が半分になるので半数体(haploid、nと表記)と言う。卵と精子が受精すると、両方の核が融合して受精卵は再び染色体がペアになった二倍体(diploid、2nと表記)となる。
減数分裂の過程は、体細胞分裂のM期の過程とよく似ているが、次に述べる4つの点で体細胞分裂と大きく異なる。
1)減数分裂の過程では、連続した2回の核および細胞質の分裂がおこり、合計4つの細胞が生じる。
2)核の分裂は2回連続しておこるが、DNA(と他の染色体の成分)は、減数分裂の最初の間期に、一度だけ複製される。
3)減数分裂により生じた4つの細胞は、一倍体の染色体、すなわち相同染色体の一方のみを含む一セットを持っている。
4)減数分裂では、両親の遺伝情報が混ぜ合わされるので、それによって生じる一倍体の細胞は、それぞれ、たった一個しかない遺伝子の組み合わせを持つようになる。
細胞内でつくられたタンパク質は、小胞体へ入るものの他にも、その使用目的に応じた適切な場所に送り込まれるために、ソートされる。たとえば、リボソームの構成要素となるタンパク質は核へ向かい、核膜孔を通って核内に入り、そこでrRNAと複合体をつくる。また、ミトコンドリアに入ってエネルギー産生を担う酵素となるものもある。ソーティングの詳しいことは、この学習の範囲を越えているので、ここでは省略する。
2)減数分裂の過程
●前期Ⅰ
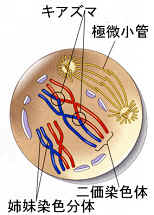
有糸分裂と同様に、減数分裂が始まる前のS期に、染色体は複製され、前期Ⅰにはセントロメアで融合した二本の染色分体からなる。
染色分体がまだ細長い間に、相同染色体同士が縦に並び始んで対合(synapsis)し、4本の染色分体が一つになる。このときの染色体を、二価染色体(vivalent or tetrad)と呼ぶ。
対合によってピッタリとくっつくので、姉妹染色分体の一方同士の間で交叉(乗り換え、crossing over)がおこる(交叉点をキアズマという)。染色分体の乗り換えによって、遺伝子の組み換え(genetic recombination)がおこる。こうして次の代に伝えられる遺伝的な変異が増大することになる。
対合や交叉のようなユニークな過程以外は、体細胞分裂前期の過程と同じである。●中期Ⅰ
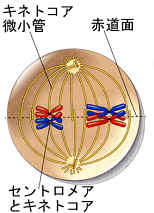
前期Ⅰは二価染色体が赤道面に並んだところで終わり、細胞は中期Ⅰに入っている。相同染色体のそれぞれのキネトコアは、紡錘糸によって一方の極だけに結び付けられているので、二本の相同染色体はそれぞれ別の極に結び付けられることになる(体細胞分裂ではこれとは対照的に、姉妹染色分体のキネトコアは両方の極に結び付けられている)。
●後期Ⅰ
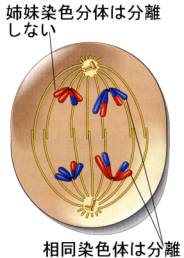
後期Ⅰでは、相同染色体は分離し、別々の極へ移動していく。各極は、父親由来と母親由来の相同染色体のどちらか一本だけをランダムに組み合わされたものを受け取る。ここでも組み合わせの違いによる変異が生じる。姉妹染色分体はまだセントロメアで結合している。
●終期Ⅰと細胞質分裂
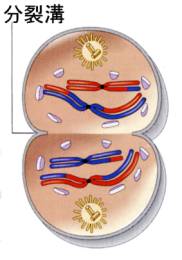
終期Ⅰでは、染色分体はいくらか脱凝縮して核膜が形成され、細胞質分裂が起きる。インターキネシスと呼ばれる間期に似た期間には、染色体の複製は行われないのでS期は存在しない。
●前期Ⅱ
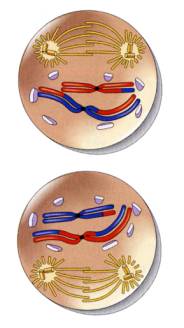
染色体は分裂の間も部分的に凝縮したままなので、二度目の減数分裂の前期もまた短い。前期Ⅱでは、多くの点で体細胞分裂の前期に似ている。相同染色体の対合は起こらず、交叉もない。
●中期Ⅱ
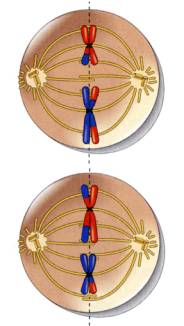
中期Ⅱでは、染色体は細胞の赤道面に並ぶ。最初の中期と二度目の中期とは簡単に区別する事ができる。中期Ⅰでは、染色分体は4本(二価染色体)に束ねられているが、中期Ⅱでは2本である。
●後期Ⅱ
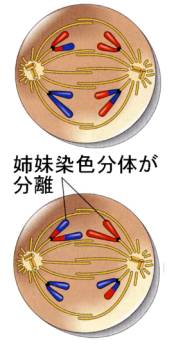
後期Ⅱでは、キネトコアで紡錘糸と結合している染色分体が、体細胞分裂の後期と同様に引き離され、反対の極へ移動していく。
●終期Ⅱと細胞質分裂
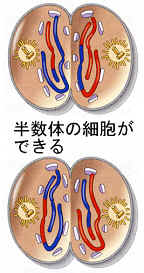
終期Ⅱでは各極には各相同染色体の対の一方が存在している。いずれも複製されていない染色体である。それから核膜が形成され、染色体は次第に伸びて染色質を形成し、細胞質分裂が起きる。
3)減数分裂による遺伝的多様性
連続2回の分裂は4つの一倍体の核をつくり、それぞれが各染色体の1本を持ち、生じる一倍体の細胞はそれぞれ異なる遺伝子の組み合わせを持つ。
この遺伝的多様性は、次の二つの理由による。
1)中期Ⅰで父方と母方の染色体が交叉によって混ぜ合わされ、まったく新しい組み合わせが生じる。
2)後期Ⅰで各対の一方が独立してランダムに分配される。
上に述べた1)と2)はまったくランダムにおこる。こうして、両親からの染色体を混ぜ合わせて次の代に伝える生殖細胞が作られる。同じ個体が作る生殖細胞でも、100個あれば100通りの異なる遺伝的組成(違いの大小はあるが)を持つことになる。
これまで述べてきたように、細胞周期の回転はきわめて重要なので、厳密に制御されている。たとえば、G1期からS期に移るときには、細胞の成長が十分であるか、複製に必要な原材料が十分あるか、などがチェックされる。またG2期からM期に移るときのも、DNAの複製は完了したか、複製の誤りはなかったか、などがチェックされる。もしもこれらの準備が十分でなければ、未熟な細胞を作ってしまったり、うまく分裂が進まない結果を生むからである。細胞周期内には、このようなチェックポイントが何ヶ所かあり、次の過程に進んでいいかどうかのチェックがされている。
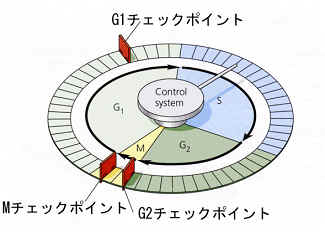
最初に見つかったチェックポイントはG2期からM期への移行に関するチェックポイントだった。ヒトデの受精卵で、受精後最初に起こる分裂に伴って特殊な因子が増加することが見つかり、卵成熟促進因子と名づけられた。酵母を使った別な研究では、細胞周期に伴って細胞の中にM期を促進する因子が増減することが見つかった。これらは同じもので、M期促進因子(M-Phase promoting factor、MPF)という名が定着する。
MPFは酵素とそれに結合するタンパク質の複合体で、両者が結合すると因子としての活性が生まれることも明らかになる。MPFが活性を持つと、直接的あるいは間接的にM期のさまざまな現象、たとえば核膜の消滅などをひきおこす。
こうしてチェックポイントというのは、酵素の活性が上がることであるということがわかってきた。G2期からM期以外のチェックポイントでも、基本的にはこれと同じようなメカニズムがはたらいている。ガン化した細胞では、これらの細胞周期チェックポイントの機能がうまくはたらかなくなっている。
http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/C/CellCycle.html#steps
![]() この章のpdfファイルをダウンロードするには、左の「Adobe」のアイコンを右クリックして、ファイルを保存を選んで、自分のパソコンにダウンロードしてください。
この章のpdfファイルをダウンロードするには、左の「Adobe」のアイコンを右クリックして、ファイルを保存を選んで、自分のパソコンにダウンロードしてください。
![]() ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。
ブロードバンド接続の場合で、ワードのファイルを望む人は、「W」のアイコンを右クリックしてください。