1.細胞膜の構造
2.お隣同士、仲よく接して
3.細胞の接着
4.外界とのインターフェイス
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
Lipid and Membrane: Biochemistry462a, University of Arizona
上記のWebページを見るためにchimeというブラウザーのプラグインをダウンロード
してインストールする事を勧めます。
更新日:2002/06/07
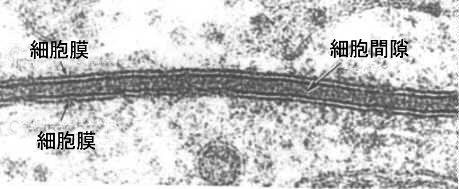
次の図は、透過型電子顕微鏡写真で撮影した、接している2つの細胞の細胞膜である。2枚の細胞膜の断面をよく見ると、電子密度が高い(黒い)二枚の膜が白い部分をはさんでいるのが見える。
電子顕微鏡の解像度をもってしても、これ以上、細胞膜の構造を明確にすることは困難であった。そこで、いろいろな知見から細胞膜の構造を示すモデルが提出された。現在、最も確からしい細胞膜の構造について説明しよう。
1)リン脂質
細胞膜の主な構成要素がリン脂質である。一つ一つのリン脂質分子は次の左図のような構造をしている。ここでは細胞膜に最も多いフォスファチジルコリンを示してある(濃い灰色:炭素、薄い灰色:水素、赤:酸素、黄色:リン)。
単純脂質である中性脂肪は、グリセロール(グリセリン)と脂肪酸のトリグリセリドだが、細胞膜を構成しているリン脂質は複合脂質で、グリセロールの3本目の水酸基は脂肪酸とではなく、リン酸とエステル結合をして、リン酸はさらに極性の高い分子(次図の場合はコリン)と結合する。
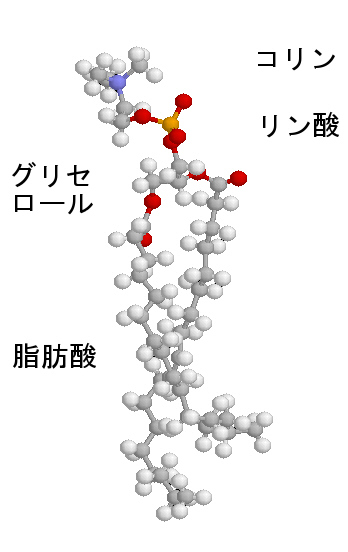
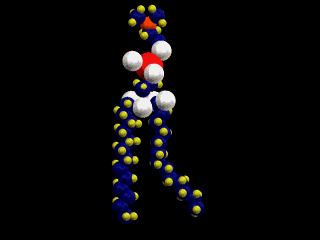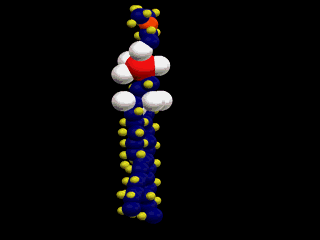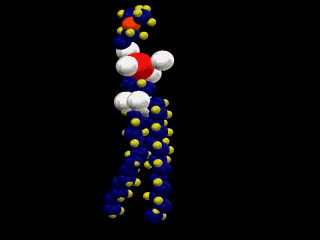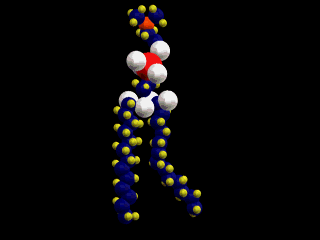
したがって、この分子は、図にあるように2本の疎水性の尾部(tail)と極性の高い頭部(head)を分子の中にあわせ持つ。脂肪酸分子中に1個の不飽和部があると、脚は二重結合のために、そこで折れ曲がるような形を取る。
2)単位膜
細胞膜は、このような分子が次の図のように配列してできているため、表面は親水性、内側は疎水性の性質を示す。このような膜を人工的にも作ることができる。
膜として安定していられるのは、水の力によるところが大きい。水(下の図の水色の部分)が、リン脂質分子のサンドイッチを離れないように押しつけているのである。
細胞膜以外にも、小胞体の膜、ミトコンドリアの内膜、外膜、ゴルジ体の膜など、細胞内に見られる膜は、基本的にはこのような構造をしていると考えられていて、単位膜(unit membrane)と呼ばれている。

3)膜タンパク質
細胞膜にはリン脂質以外にも多くの分子が存在する。タンパク質やそれに付随する構造を書き入れた模式図が次の図である。細胞膜に埋め込まれたタンパク質を、膜タンパク質(membrane protein)と呼ぶ。
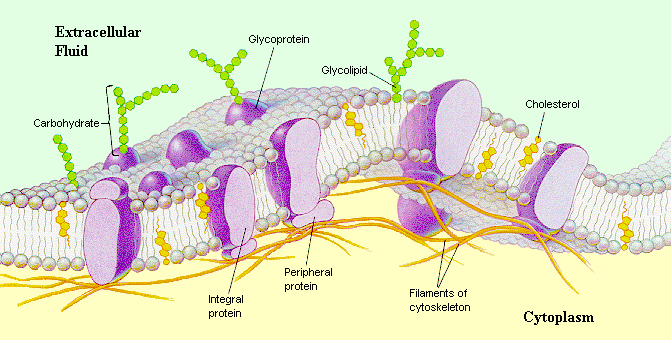
この図に見られるように、細胞膜は脂質の二重膜(lipid bilayer)の海に、膜タンパク質が氷山のように頭を少し出して浮かんだような構造をしていると考えられている。この構造モデルを流動モザイクモデル(fluid mosaic model)と呼んでいる。
脂質の二重膜は、膜タンパク質を浮かべる担体のはたらきをしているだけで、極言すれば細胞膜の性質は、膜タンパク質の性質に依存している。脂質の二重膜の海と書いたように細胞膜には流動性があり、膜タンパク質はこの海の中を自由に動くことができる。
ただし、上の図にperipheral proteinと書かれたようなタンパク質が動きを止める場合もあるし、後で述べる密着結合を越えて動くことはできない。
たとえば、脂質の二重膜は、上に述べたような分子の配列のために、きわめて小さな分子か疎水性の炭化水素のような分子は通すが、大部分の分子やイオンを通さない(次表)。
|
分子の性質 |
例 |
透過性 |
| 疎水性分子 | N2、O2、炭化水素 | 自由に透過 |
| 極性のある小分子 | H2O、CO2、グリセロール、尿素 | 自由に透過 |
| 極性のある大分子 | ブドウ糖などの単糖類、二糖類 | 透過できない |
| イオンや電荷を持つ分子 | アミノ酸、H+、HCO3-、Na+、K+、Ca2+、Cl-、Mg2+ | 透過できない |
溶媒のような小さな分子は通すが、溶質は通さない性質を持つ、このような膜を半透膜 (semipermeable membrane)という。しかしながら、この性質だけでは細胞は必要な物質を取り込むことができない。
そこで膜タンパク質の登場である。膜タンパク質の中には、膜を貫通しているものがある。多くの膜貫通タンパク質は、ちょうど弁の付いた管のようになっていて、特定のイオンや物質を選択的に通すことができる。このタンパク質のおかげで、細胞膜は特定のイオンやブドウ糖などの必要な物質を通すことができるのである。
したがって細胞膜は、選択的透過性を示す半透膜ということができる。すべての細胞に同じ膜タンパク質が埋め込まれているわけではないので、どんな物質が通過できるかは細胞によって異なる。
細胞膜は物質の出入りをコントロールするばかりでなく、隣の細胞と接するためのインターフェイスとなっている。まずは、細胞どうしの結びつき合いから話を進めて行こう。
関連するサイトとリンク(このページへ戻るときはブラウザーの戻るを選んでください)
TRANSPORT
IN AND OUT OF CELLS
すでに話したように、単細胞生物を除いて、細胞は孤立して存在することはない。お隣同士仲よく接して組織を作り上げている。たとえば上皮細胞なら、ふつうは上皮組織の中で周囲と協調しつつ、上皮組織の一員としての働きをしている。もしも勝手な行動を始めたらそれはガン細胞である。
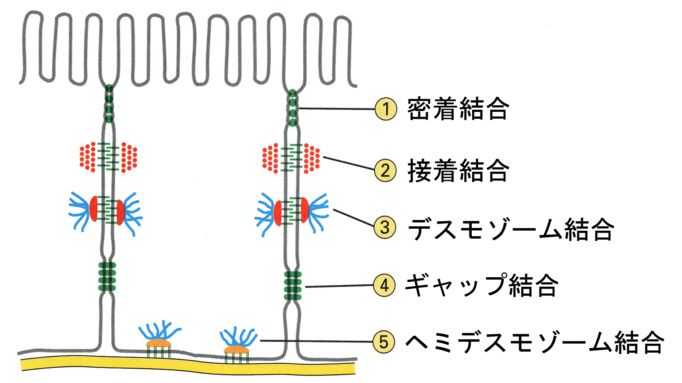
このような場合、細胞どうしは特定の構造で結合しあっている。その構造には大きく分け て4つある。
①密着結合(tight junction)
②接着結合(adherence junction)
③デスモゾーム結合(desmosome
junction)と⑤ヘミデスモゾーム
④ギャップ結合(gap junction)
である。
1)密着結合(タイトジャンクション)
タイトジャンクションは、その名前が示すとおりがっちりした結合である。下の模式図にあるように、タイトジャンクションタンパク質はジッパーのように隣り合った細胞の細胞膜を連続的につなぎあわせる。
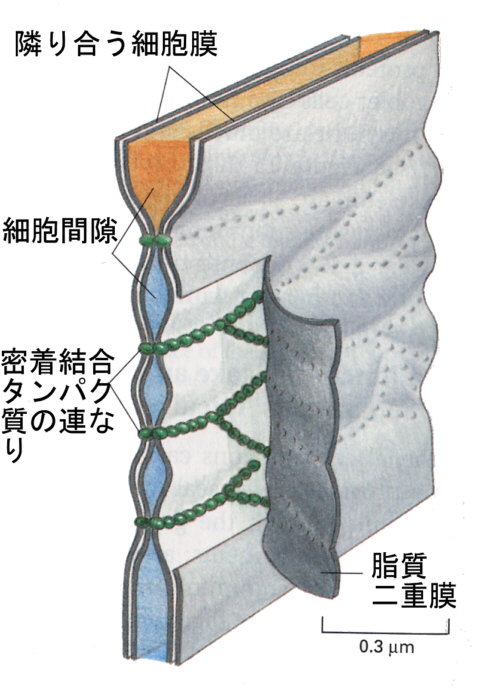
そのために、細胞間隙(intercellular space)と細胞の自由表面は不連続になる。このタイトジャンクションのために、上皮細胞のシートは外側と内側(腸上皮組織の場合でいえば管腔側と組織内)を分けるバリヤーになり、溶質は自由に組織の内側に入ることことができない。
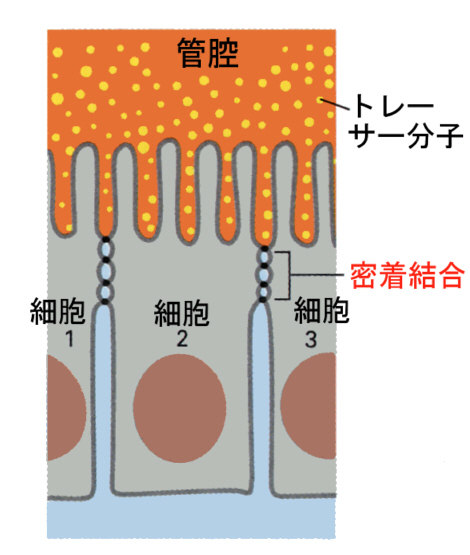
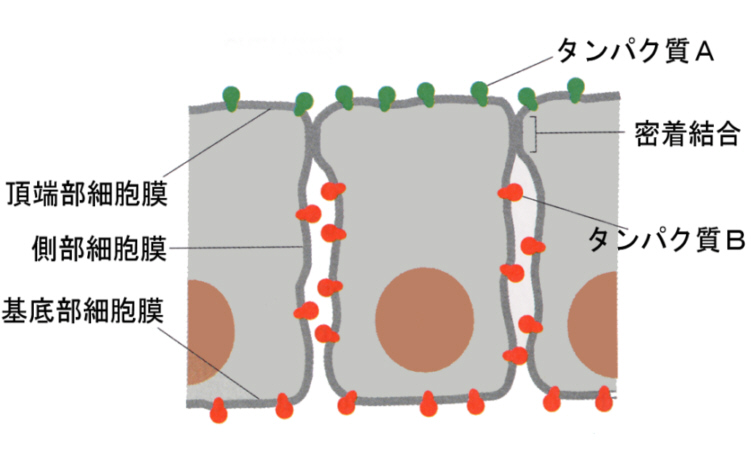
また、タイトジャンクションは膜タンパク質の自由な拡散を妨げるので、タイトジャンクションで分けられた二つのコンパートメント(細胞膜の区画)の膜タンパク質は混じり合うことができなくなる。下の図は、細胞の頂端部(apical)細胞膜の膜タンパク質Aと基底膜側(basal)の膜タンパク質Bが、密着結合によって自由に混じり合わないことを示す模式図である。
このため、両者の膜タンパク質の性質が異なれば、溶質の細胞通過に方向性が生まれることになる。
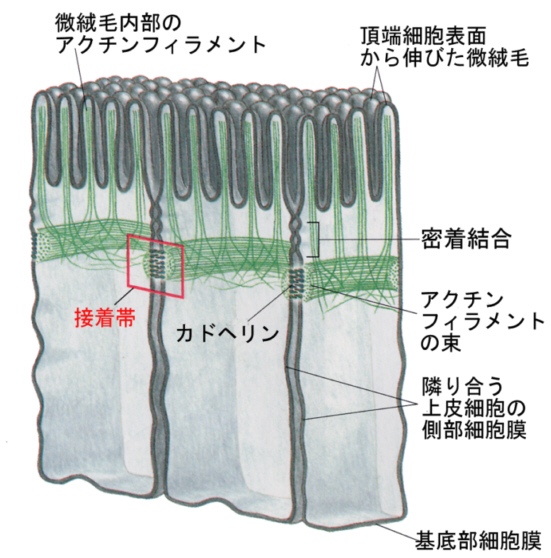
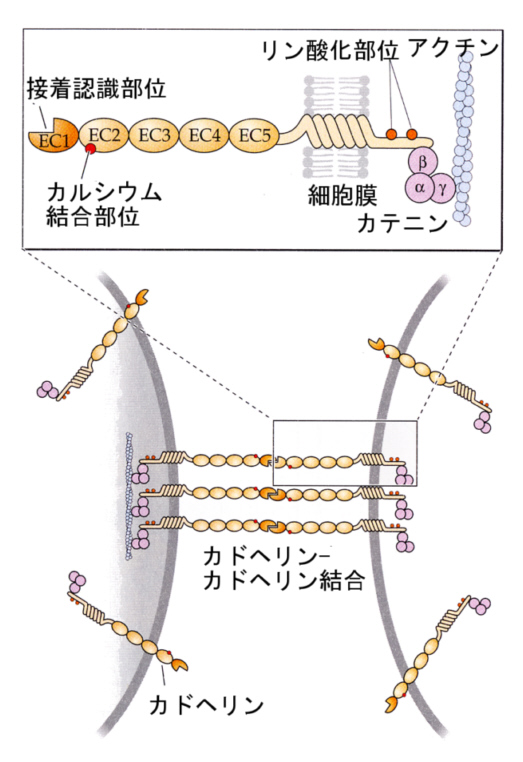
2)接着結合
接着結合は、タイトジャンクションの下に帯のように細胞周囲にあって、接着帯を構成している。次に述べる デスモゾームと似ているがボタン状タンパク質がなく、カドヘリン(後述)という膜タンパク質どうしが接着して、細胞をつなぎ止めている。
カドヘリンの細胞内の端は、細胞骨格の一つであるアクチンフィラメントの束と結合している。
3)デスモゾーム結合
次の電子顕微鏡写真は、イモリの表皮に見られるデスモゾームである。
デスモゾームは、ワイシャツかブラウスのちからボタンのような円盤状のタンパク質と、細胞の外側に向かって細胞膜を貫通する結合タンパク質から構成されている。この結合タンパク質もカドヘリンである。
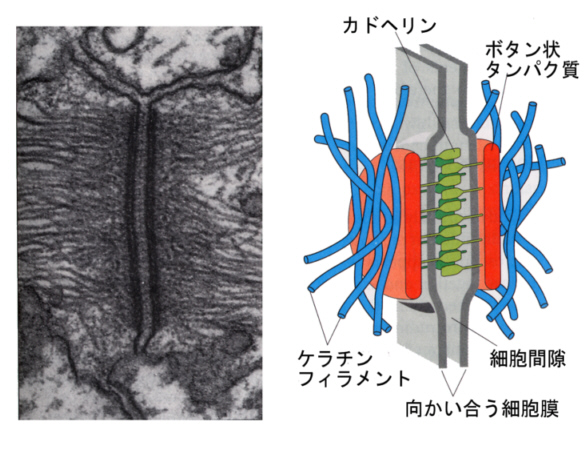
ボタン状タンパク質は細胞膜のすぐ内側にあって、ここから突き出す結合タンパク質が、隣の細胞のデスモゾームの結合タンパク質と結合している。ちょうど2枚の布を2個のボタンを介して糸で綴じ合わせたような構造をしている。ボタン状タンパク質には中間径フィラメントのケラチンフィラメントが結合している。
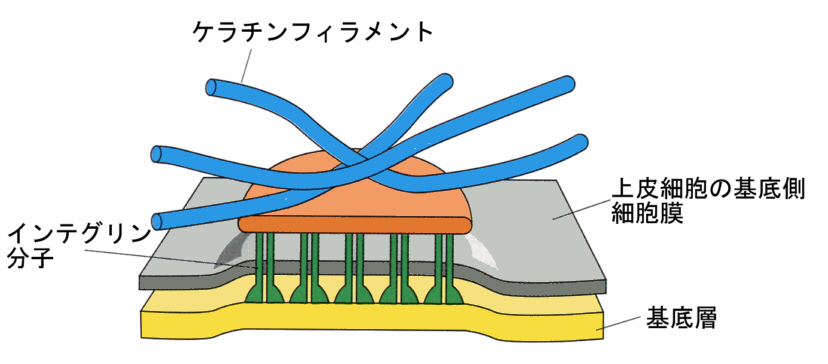
上皮細胞の底にも同じ構造があるが、結合タンパク質は隣の細胞のデスモゾームではなく、基底層(basal lamina)と直接、結合している。この構造をヘミデスモゾーム(hemi=半分)と呼ぶ。
4)ギャップ結合
ギャップジャンクションは、下の図にあるように、管状の膜貫通タンパク質が隣の細胞のものと結びついた構造をしている。このため、2つの細胞の細胞質は連続することになる。
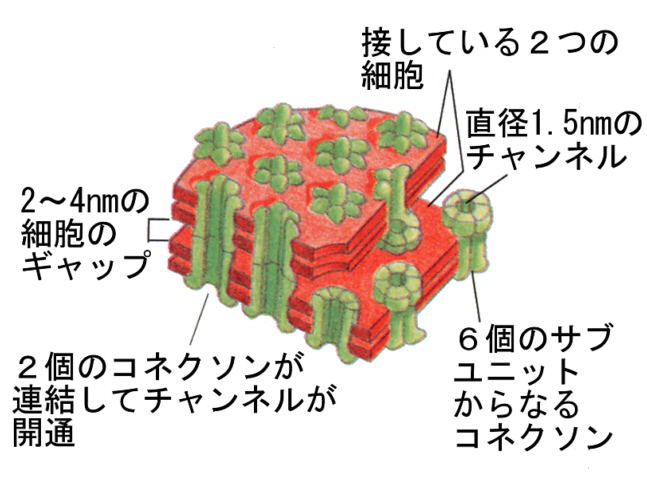
ギャップジャンクションの穴(1.5 nm)は、分子量1000以下の分子を通すことができる。穴はサイトゾールのカルシウムイオンの濃度によって開閉する(低いと開き、高いと閉じる)。ギャップジャンクションによって、細胞どうしが電気的につながったり、カルシウムイオンのような小さい分子を通過させることにより細胞間の同調が行われていると考えられている。
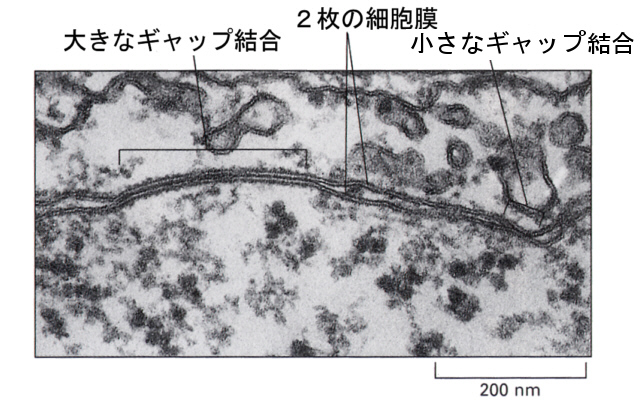
次の図の電子顕微鏡写真では、細胞膜の中ほどにある面積の大きなギャップジャンクションと、右の方に小さなギャップジャンクションが見える。
小腸上皮組織は、これまで述べたような結合と膜タンパク質の性質によって、一枚の上皮細胞のシートを形成し、個々の細胞ではなくシートとして機能している。
1)接着の過程
上に述べたような細胞どうしの結合が作られるためには、まず最初に細胞どうしが接着(adhere)しなければならない。細胞はただくっつきあっているのではなく、選択的な接着によって、積極的に寄り集まっているのである。
接着の過程は、
1)細胞がお互いに相手の細胞を認識して、同じ仲間ならくっつきあう
2)同じ仲間同士が細胞の塊を作る
3)細胞の塊はやがて分化して組織となる
というように進んで行く。
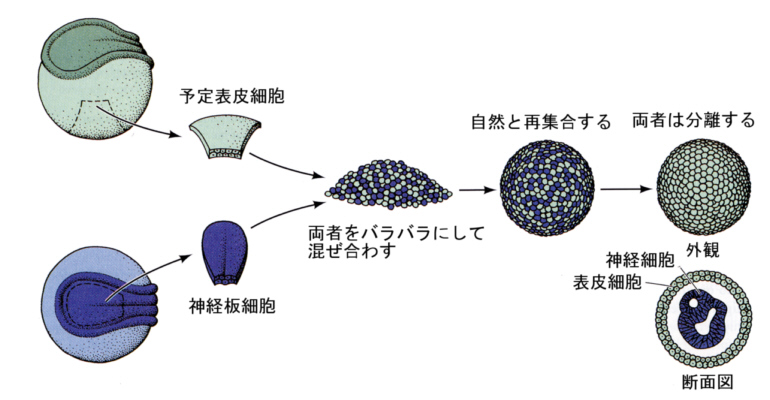
次の図は、イモリの予定表皮細胞と神経板の細胞を混ぜ合わせておくと、自然と両者は塊を作り、やがて選別が起こり表皮は外側に、神経細胞は内側に塊を作り、それぞれの組織に分化することを示した、ホルトフレーターらの実験である(1955)。この古典的な実験は、細胞はお互いに認識しあい選別が起こることを示している。
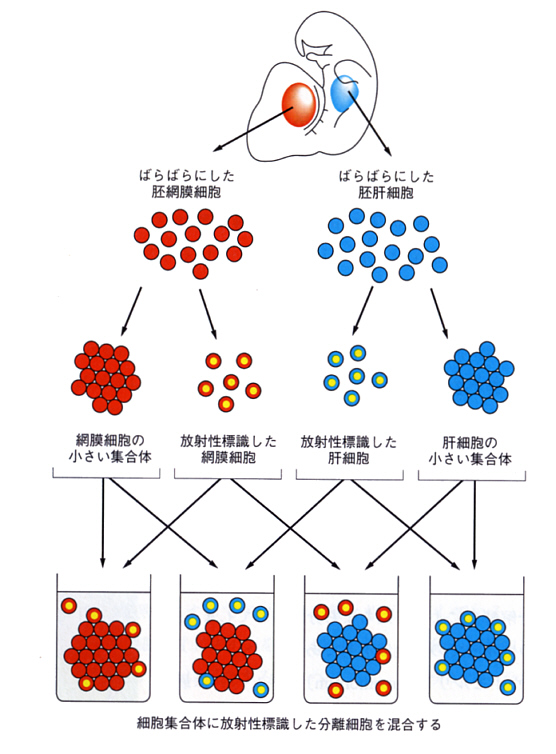
次の図は、胚(embryo)からとった網膜(retina)と肝臓の細胞をばらばらにして、混ぜ合わせた時の様子を、一部の細胞に放射性同位元素(radioactive isotope)で標識して調べたときの模式図である。時間を追って細胞塊の放射能を計測すると、網膜細胞あるいは肝臓細胞どうしは細胞塊を作るが、違った細胞はほとんど混ざらないことを定量的に示すことができる。
このような細胞の選択的な接着過程は、カイメンのような系統的に古い動物でも観察することができる。磯によく見られるダイダイイソカイメンとムラサキソカイメンはきわめて近縁種で体の色が異なるが、これを、それぞればらばらにして混ぜておくと、最初は混ざっているが、やがてもとの2つの色の塊に分かれてくる。
このような過程は、発生(development)の進行や形態形成(morphogenesis)に伴ってごく普通に起こる現象である。発生や形態形成の過程では、それまで発現していた接着分子が消失し、別の接着分子が発現して細胞膜の接着性が変わり、細胞の選別が行われる。
2)接着分子
それでは、接着はどのようにして起こるのだろうか。細胞どうしが相互に認識して接着するためには、細胞膜の表面に突き出した接着タンパク質が関与することが分かってきた。主な接着分子には、カドヘリン(Ca+adhere+in)とCAM(cell adhesion molecule)がある。いずれの場合も、接着分子どうしが、結合して細胞を接着させる。
カドヘリンは名前が示すとおり、カルシウム依存性の接着分子である。カルシウムイオンの存在下に、隣あった細胞のカドヘリンどうしが、お互いに同じ分子だということを認識して結合する(これhomophilicという)。上皮細胞だと、こうして形成された接着がジャンクション(adhesion belt, desmosome)になる。カドヘリンにはいくつかの種類があって、発生(development)の際の分化(differentiation)に重要な役割を演じている。
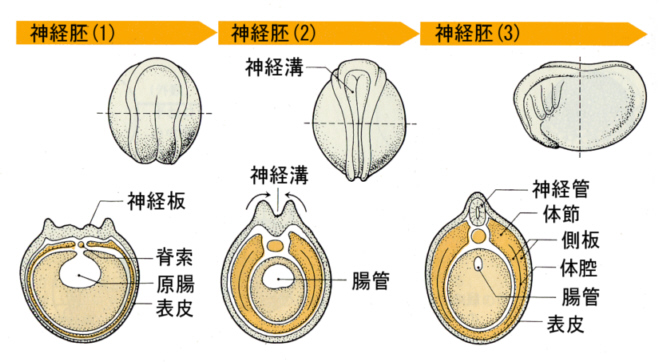
次の図は、カエルの初期発生において、嚢胚期から神経胚期へ進んだあと、外胚葉に神経板(neural plate)ができ、やがて左右の縁が盛り上がって中央で閉じて神経管(neural tube)をつくる過程を描いている。さらに神経管と表皮の間に神経冠(neural crest)が分かれ出て、移動して神経節(ganglion)を生じる。
この過程で起こる細胞の選別の過程は、次のようなメカニズムによることが分かってきた。
次の図は、マウスのカドヘリンのうち、表皮に発現するE-カドヘリンと、神経組織に発現するN-カドヘリンの発現の消長によって、神経管が分化することを示す図である。
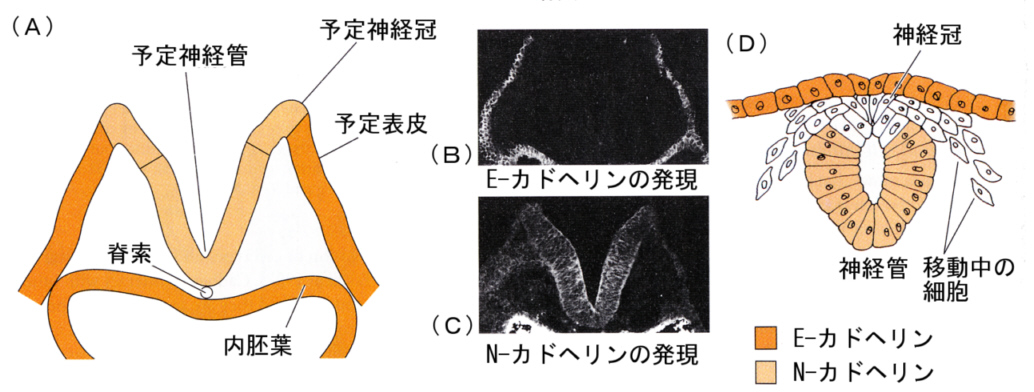
初めは表皮であった細胞ではE-カドヘリンが発現しているが、やがて将来、神経管になる部域ではE-カドヘリンが発現しなくなり、かわりにN-カドヘリンガ発現してくる。(B)と(C)の蛍光顕微鏡写真は、同じ切片を蛍光色素を変えて示したものである。
N-カドヘリンを発現する細胞群はE-カドヘリンを発現している細胞とは一緒にいられなくなり、そこから内側にくびれ出て、神経管を形成する。
神経冠細胞になる細胞はどちらも発現せず、そのため表皮と神経管の間に両者から別れ出て、神経管を覆うように分布する。やがて神経冠細胞は移動をはじめる。
カドヘリンは、細胞内でカテニンというタンパク質を介して、細胞骨格分子であるアクチン繊維と結合している。
CAMはカドヘリンとは異なり、カルシュウム非依存性の接着分子である。やはり隣の細胞のCAMと結合する(異なる膜タンパク質と結合することもあるが)。CAMには、いろいろな種類があり、癌の転移や、発生に伴う細胞のソーティングに関係するらしいが、詳しいことは省略する。
CAMは、後で免疫の話をするときに登場する、免疫グロブリン(immunogloburin, Ig)と似た構造をしていて、免疫グロブリンスーパーファミリーの一員である。
さらに学びたい人へ(このページへ戻るときはブラウザーの戻るを選んでください)
http://www.neuro.wustl.edu/neuromuscular/lab/adhesion.htm
3)細胞外マトリックスとの接着
上皮細胞のシートは、結合組織の一番表面の基底層(basal lamina)に張りついている。この張りつき(ジャンクション)は、すでに述べたように構造としてはヘミデスモゾームによる。ヘミデスモゾームの場合、膜タンパク質として細胞表面へ突き出しているのは、インテグリンと呼ばれるタンパク質である。インテグリンは、基底膜を構成するラミニンやフィブロネクチン、コラーゲンでできた網目構造にしっかりと結合している。
前のセクションではジャンクションと接着に関連して、いくつかの膜タンパク質を紹介した。細胞膜にはさらに多くの種類の膜タンパク質が埋め込まれていて、これらが外界との機能的なインターフェイスの役割を担っている。
主なものに、1)輸送の役割を担うタンパク質、2)受容体の役割を担うタンパク質、がある。輸送と受容体の両方の役割をするタンパク質もある。
1)膜内輸送タンパク質(membrane transport protein)
すでに、この節の冒頭で述べたように、細胞膜は選択的透過性を示す半透膜である。この選択的透過性は、膜に埋め込まれた膜内輸送タンパク質を通して、決まったモノのみが移動するからである。
膜内輸送タンパク質には、大きく分けて(1)チャンネルタンパク質と(2)運搬タンパク 質がある。
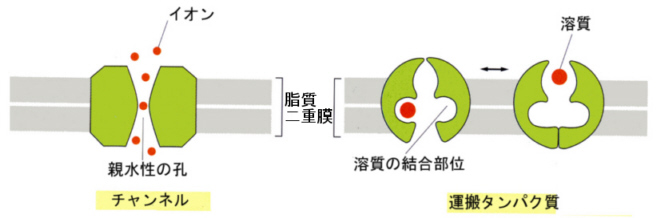
チャンネルタンパク質は、弁(ゲート)のついた管のようなもので、弁が開くと濃度勾配にしたがって、特定のイオンがチャンネルの中を通過できる。チャンネルは、脂質の二重膜を貫通する何本かのαヘリックスの棒が束ねられたような構造をしている。内径や内側に配列しているアミノ酸の側鎖の性質によって、チャンネルがどんなイオンを通せるかが決まってくる(チャンネルフィルターの存在)。
ナトリウムチャンネルやカルシュウムチャンネルがよく知られている。ゲートが開くメカニズムには、電位変化による方式と、受容体に情報分子が受容される方式の2つがある。
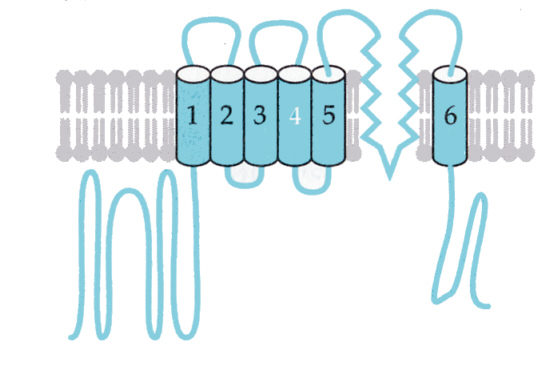
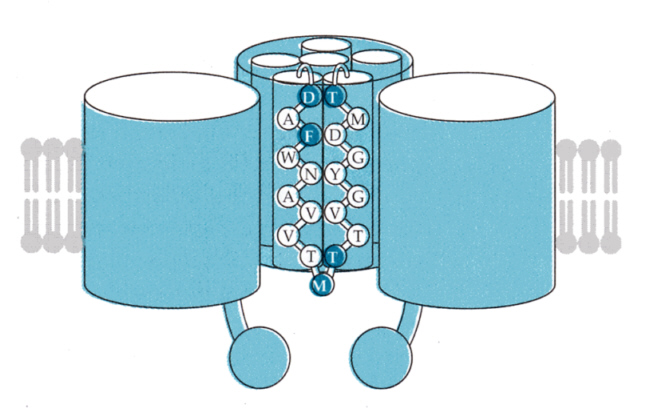
次の図は、電位型のカリウムチャンネルの模式図である。6本のαヘリックスと逆平行のβシートからなるユニットが4つで、1つのチャンネルを構成していると考えられている。下の図は手前の1つのユニットは描かれていない。
パワーポイントのファイルでカリウムチャンネルタンパク質の働きを解説
http://tedlab.urmc.rochester.edu/HSF-01/HSF01-Na%20&%20K%20Channels.ppt
運搬タンパク質は、分子内に溶質が結合できる場所があり、上の図にあるように、この結合部位を介して溶質を通過させることができる。濃度勾配にしたがって溶質が通過する場合もあるが、ATPのエネルギーを使って濃度勾配に逆らって溶質を輸送する場合もある。
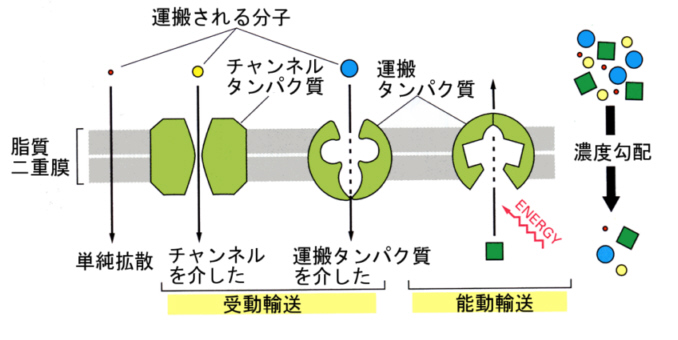
細胞外へナトリウムイオンを汲み出し、カリウムイオンを汲み入れる働きをする膜タンパク質がある。この膜タンパク質は、Na+-K+ ATPaseと呼ばれることも、ナトリウムポンプと呼ばれることもある。エネルギーを使う、このような輸送を能動輸送(active transport)と呼んでいる。
2)受容体タンパク質(receptor protein)
白血球の一つであるマクロファージ(macrophage、大食細胞あるいは貪食細胞と訳す、macro=大きい、phago=食べる)は、固形物を囲み込んで細胞内へ取り込み、消化することができる(ファゴサイトーシスphagocytosis、食細胞運動という)。
もっと小さな分子を選択的に取り込むためには、細胞膜表面に突き出した受容体により選択的に捕捉した後、エンドサイトーシスによる取り込みが行われる。
3)情報を受信するアンテナとしての受容体
細胞の外から作用する情報分子で細胞膜を通過できないものは、細胞表面にある受容体と結合してその作用をあらわす。ホルモンや神経伝達物質は受容体と結合し、つぎにトランスデューサーとして働く別の膜タンパク質にその情報が伝わり、最後に細胞内情報伝達系に情報が伝えられる。
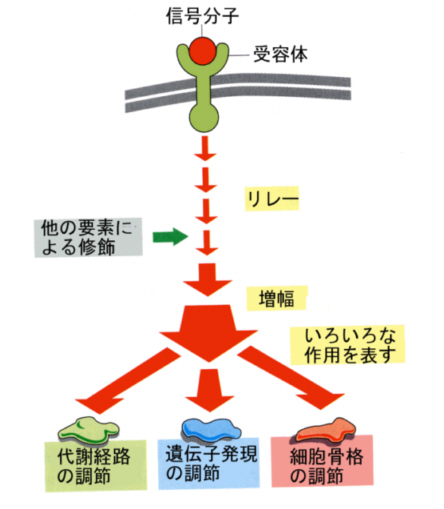
この点に関しては、後で再び学ぶ。
![]() この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。
この章のpdfファイルをダウンロードするには、左のアイコンを右クリックしてください。